Unique Features of Entamoeba Sulfur Metabolism; Compartmentalization, Physiological Roles of Terminal Products, Evolution and Pharmaceutical Exploitation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Protozoologica Special Issue: Protists in Soil Processes
Acta Protozool. (2012) 51: 201–208 http://www.eko.uj.edu.pl/ap ActA doi:10.4467/16890027AP.12.016.0762 Protozoologica Special issue: Protists in Soil Processes Review paper Ecology of Soil Eumycetozoans Steven L. STEPHENSON1 and Alan FEEST2 1Department of Biological Sciences, University of Arkansas, Fayetteville, Arkansas, USA; 2Institute of Advanced Studies, University of Bristol and Ecosulis ltd., Newton St Loe, Bath, United Kingdom Abstract. Eumycetozoans, commonly referred to as slime moulds, are common to abundant organisms in soils. Three groups of slime moulds (myxogastrids, dictyostelids and protostelids) are recognized, and the first two of these are among the most important bacterivores in the soil microhabitat. The purpose of this paper is first to provide a brief description of all three groups and then to review what is known about their distribution and ecology in soils. Key words: Amoebae, bacterivores, dictyostelids, myxogastrids, protostelids. INTRODUCTION that they are amoebozoans and not fungi (Bapteste et al. 2002, Yoon et al. 2008, Baudalf 2008). Three groups of slime moulds (myxogastrids, dic- One of the idiosyncratic branches of the eukary- tyostelids and protostelids) are recognized (Olive 1970, otic tree of life consists of an assemblage of amoe- 1975). Members of the three groups exhibit consider- boid protists referred to as the supergroup Amoebozoa able diversity in the type of aerial spore-bearing struc- (Fiore-Donno et al. 2010). The most diverse members tures produced, which can range from exceedingly of the Amoebozoa are the eumycetozoans, common- small examples (most protostelids) with only a single ly referred to as slime moulds. Since their discovery, spore to the very largest examples (certain myxogas- slime moulds have been variously classified as plants, trids) that contain many millions of spores. -

An Unusual Case of Amoebic Liver Abscess Presenting with Hepatic Encephalopathy: a Case Report
Case Report An Unusual Case of Amoebic Liver Abscess Presenting with Hepatic Encephalopathy: A Case Report Anil Kumar SARDA, Rakesh MITTAL Submitted: 16 Sep 2010 Department of Surgery, Maulana Azad Medical College and Lok Nayak Accepted: 3 Jan 2011 Hospital, New Delhi 110 002, India Abstract Amoebic liver abscess (ALA) with jaundice and encephalopathy is a rare occurrence and has been recognised and studied more frequently in recent years. We present a case of massive ALA presenting with jaundice, hepatic encephalopathy, and septicaemia that was treated successfully with percutaneous drainage of the abscess, right-sided chest tube insertion, and anti-amoebic therapy. Keywords: amoebiasis, hepatic encephalopathy, hepatology, jaundice, liver abscess Introduction On chest examination, there was bilateral equal air entry. Upon investigation, haemoglobin was Amoebic liver abscess (ALA) is the most 11 g/dL, with a total leukocyte count of frequent extra-intestinal manifestation of 13 000 cells/mm3 (normal range is 4000– Entamoeba histolytica infection. It has been 11 000 cells/mm3). Liver function tests revealed reported that jaundice is uncommon and mild total serum bilirubin of 20 mg/dL, with direct in liver abscess, and some even consider the bilirubin of 15 mg/dL, serum glutamic-oxaloacetic presence of jaundice as a feature against the transaminase (SGOT) of 324 IU/L (normal level diagnosis of hepatic amoebiasis (1). The cause of is less than 40 IU/L), serum glutamic–pyruvic jaundice in a case of ALA has been hypothesised transaminase (SGPT) of 340 IU/L (normal level to result from either hepatocellular dysfunction or is less than 40 IU/L), and alkaline phosphatase intrahepatic biliary obstruction (2). -

Comparative Genomics of the Social Amoebae Dictyostelium Discoideum
Sucgang et al. Genome Biology 2011, 12:R20 http://genomebiology.com/2011/12/2/R20 RESEARCH Open Access Comparative genomics of the social amoebae Dictyostelium discoideum and Dictyostelium purpureum Richard Sucgang1†, Alan Kuo2†, Xiangjun Tian3†, William Salerno1†, Anup Parikh4, Christa L Feasley5, Eileen Dalin2, Hank Tu2, Eryong Huang4, Kerrie Barry2, Erika Lindquist2, Harris Shapiro2, David Bruce2, Jeremy Schmutz2, Asaf Salamov2, Petra Fey6, Pascale Gaudet6, Christophe Anjard7, M Madan Babu8, Siddhartha Basu6, Yulia Bushmanova6, Hanke van der Wel5, Mariko Katoh-Kurasawa4, Christopher Dinh1, Pedro M Coutinho9, Tamao Saito10, Marek Elias11, Pauline Schaap12, Robert R Kay8, Bernard Henrissat9, Ludwig Eichinger13, Francisco Rivero14, Nicholas H Putnam3, Christopher M West5, William F Loomis7, Rex L Chisholm6, Gad Shaulsky3,4, Joan E Strassmann3, David C Queller3, Adam Kuspa1,3,4* and Igor V Grigoriev2 Abstract Background: The social amoebae (Dictyostelia) are a diverse group of Amoebozoa that achieve multicellularity by aggregation and undergo morphogenesis into fruiting bodies with terminally differentiated spores and stalk cells. There are four groups of dictyostelids, with the most derived being a group that contains the model species Dictyostelium discoideum. Results: We have produced a draft genome sequence of another group dictyostelid, Dictyostelium purpureum, and compare it to the D. discoideum genome. The assembly (8.41 × coverage) comprises 799 scaffolds totaling 33.0 Mb, comparable to the D. discoideum genome size. Sequence comparisons suggest that these two dictyostelids shared a common ancestor approximately 400 million years ago. In spite of this divergence, most orthologs reside in small clusters of conserved synteny. Comparative analyses revealed a core set of orthologous genes that illuminate dictyostelid physiology, as well as differences in gene family content. -

Amebiasis Annual Report 2017
Amebiasis Annual Report 2017 Amebiasis Amebiasis is no longer a reportable disease in Louisiana. Outbreaks, however, should still be reported. Amebiasis (amoebiasis) is a parasitic infection caused by Entamoeba histolytica or Entamoeba dispar. The parasite is transmitted by the fecal-oral route, either through direct contact with feces or through the consumption of contaminated food or water. Between 80% and 90% of infected individuals develop no symptoms. For symptomatic cases, the incubation period between infection and illness can range from days to weeks. The symptoms are typically gastrointestinal issues, such as diarrhea or stomach pains. It is also possible for the parasite to spread to the liver and cause abscesses. Entamoeba histolytica can be found world-wide, but is more prevalent in tropical regions with poor sanitary conditions. In some areas with extremely adverse conditions, the prevalence can be as high as 50% in the population. There are no recent data on prevalence of amebiasis in the U.S. however, prevalence is estimated to be between 1% and 4% of the population. High risk groups are refugees, recent immigrants, travelers (particularly those who have spent long periods of time in an endemic area), institutionalized people (particularly developmentally or mentally-impaired people), and men who have sex with men. The number of cases reported within Louisiana is usually low. There are typically less than ten cases per year with a few exceptions (Figure 1). Figure 1: Amebiasis cases - Louisiana, 1970-2017 40 35 30 25 20 15 Number of Cases Number 10 5 0 70 72 74 76 78 80 82 84 86 88 90 92 94 96 98 00 02 04 06 08 10 12 14 16 Year Louisiana Office of Public Health – Infectious Disease Epidemiology Section Page 1 of 4 Amebiasis Annual Report 2017 Hospitalization Hospitalization surveillance is based on Louisiana Hospital Inpatient Discharge Data (LaHIDD). -

The Intestinal Protozoa
The Intestinal Protozoa A. Introduction 1. The Phylum Protozoa is classified into four major subdivisions according to the methods of locomotion and reproduction. a. The amoebae (Superclass Sarcodina, Class Rhizopodea move by means of pseudopodia and reproduce exclusively by asexual binary division. b. The flagellates (Superclass Mastigophora, Class Zoomasitgophorea) typically move by long, whiplike flagella and reproduce by binary fission. c. The ciliates (Subphylum Ciliophora, Class Ciliata) are propelled by rows of cilia that beat with a synchronized wavelike motion. d. The sporozoans (Subphylum Sporozoa) lack specialized organelles of motility but have a unique type of life cycle, alternating between sexual and asexual reproductive cycles (alternation of generations). e. Number of species - there are about 45,000 protozoan species; around 8000 are parasitic, and around 25 species are important to humans. 2. Diagnosis - must learn to differentiate between the harmless and the medically important. This is most often based upon the morphology of respective organisms. 3. Transmission - mostly person-to-person, via fecal-oral route; fecally contaminated food or water important (organisms remain viable for around 30 days in cool moist environment with few bacteria; other means of transmission include sexual, insects, animals (zoonoses). B. Structures 1. trophozoite - the motile vegetative stage; multiplies via binary fission; colonizes host. 2. cyst - the inactive, non-motile, infective stage; survives the environment due to the presence of a cyst wall. 3. nuclear structure - important in the identification of organisms and species differentiation. 4. diagnostic features a. size - helpful in identifying organisms; must have calibrated objectives on the microscope in order to measure accurately. -

The Nutrition and Food Web Archive Medical Terminology Book
The Nutrition and Food Web Archive Medical Terminology Book www.nafwa. -

Surveillance Study of Acute Gastroenteritis Etiologies in Hospitalized Children in South Lebanon (SAGE Study)
pISSN: 2234-8646 eISSN: 2234-8840 https://doi.org/10.5223/pghn.2018.21.3.176 Pediatr Gastroenterol Hepatol Nutr 2018 July 21(3):176-183 Original Article PGHN Surveillance Study of Acute Gastroenteritis Etiologies in Hospitalized Children in South Lebanon (SAGE study) Ghassan Ghssein, Ali Salami, Lamis Salloum, Pia Chedid*, Wissam H Joumaa, and Hadi Fakih† Rammal Hassan Rammal Research Laboratory, Physio-toxicity (PhyTox) Research Group, Lebanese University, Faculty of Sciences (V), Nabatieh, *Department of Medical Laboratory Sciences, Faculty of Health Sciences, University of Balamand, †Department of Pediatrics, Faculty of Medical Sciences, Lebanese University, Beirut, Lebanon Purpose: Acute gastroenteritis (AGE) is a major cause of morbidity and remains a major cause of hospitalization. Following the Syrian refugee crisis and insufficient clean water in the region, this study reviews the etiological and epidemiological data in Lebanon. Methods: We prospectively analyzed demographic, clinical and routine laboratory data of 198 children from the age of 1 month to 10 years old who were admitted with the diagnosis of AGE to a private tertiary care hospital located in the district of Nabatieh in south Lebanon. Results: Males had a higher incidence of AGE (57.1%). Pathogens were detected in 57.6% (n=114) of admitted pa- tients, among them single pathogens were found in 51.0% (n=101) of cases that consisted of: Entamoeba histolytica 26.3% (n=52), rotavirus 18.7% (n=37), adenovirus 6.1% (n=12) and mixed co-pathogens found in 6.6% (n=13). Breast-fed children were significantly less prone to rotavirus (p=0.041). Moreover, children who had received the rota- virus vaccine were significantly less prone to rotavirus (p=0.032). -
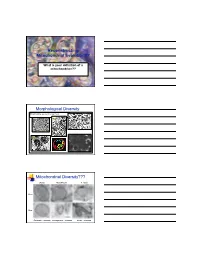
Reconstructing Mitochondrial Evolution?? Morphological Diversity Mitochondrial Diversity???
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial Diversity??? Chicken Neocallimastix T. foetus 100 nm 50 nm Entamoeba - mitosome microsporidian - mitosome Giardia - mitosome Reconstructing Evolution Mitochondrial evolution well established endosymbiotic theory α-proteobacterium - Rickettsia prowazekii Hydrogenosomal evolution No DNA NOW 2 examples Nyctotherus and Blastocystis (MLO) Several proteins similar to mitochondria Mitosome evolution No DNA Few proteins identified similar to mitochondria Origins via Endosymbiosis Current dogma - mitochondria and related Aerobic α-proteobacterium prokaryote gave rise to present day mitochondria. organelles arose just once in evolution Are hydrogenosomes and mitosomes of anaerobic protists derived from the same proto-mitochondrion? Evidence for: accumulating evidence for several proteins that are currently found in mitochondria - Proteins of Fe-S cluster formation. Scenario A Common ancestor facultative Scenario B organism? degenerate mitochondrion invoke lateral gene transfer from anaerobic prokaryotes Hypotheses for Mito Acquisition Mitochondrion-related Organelles 2 way to interpret this summary - can you think of both? Groups with mitochondrial homologues Hjort et al Phil. Trans. R. Soc. B 2010 365, 713-727 Origin of Hydrogenosomes (Mitosomes?) 1. a)Conversion of Mitochondria HYD b)Common ancestor with mitochondria ? HYD 2. Independent Origin α-proteobacterium ? - anaerobic bacterium HYD Organelles - origins and biogenesis Approaches: (1) Conduct phylogenetic analyses of similar proteins Hsp70 Fd Hsp60 Isc subunits (2) Examine protein targeting to the organelle matrix protein targeting membrane protein targeting (3) Characterize membrane/translocation components These components could have evolved as the endosymbiont was converted to organelle. Reveals evolutionary history. -

Protist Phylogeny and the High-Level Classification of Protozoa
Europ. J. Protistol. 39, 338–348 (2003) © Urban & Fischer Verlag http://www.urbanfischer.de/journals/ejp Protist phylogeny and the high-level classification of Protozoa Thomas Cavalier-Smith Department of Zoology, University of Oxford, South Parks Road, Oxford, OX1 3PS, UK; E-mail: [email protected] Received 1 September 2003; 29 September 2003. Accepted: 29 September 2003 Protist large-scale phylogeny is briefly reviewed and a revised higher classification of the kingdom Pro- tozoa into 11 phyla presented. Complementary gene fusions reveal a fundamental bifurcation among eu- karyotes between two major clades: the ancestrally uniciliate (often unicentriolar) unikonts and the an- cestrally biciliate bikonts, which undergo ciliary transformation by converting a younger anterior cilium into a dissimilar older posterior cilium. Unikonts comprise the ancestrally unikont protozoan phylum Amoebozoa and the opisthokonts (kingdom Animalia, phylum Choanozoa, their sisters or ancestors; and kingdom Fungi). They share a derived triple-gene fusion, absent from bikonts. Bikonts contrastingly share a derived gene fusion between dihydrofolate reductase and thymidylate synthase and include plants and all other protists, comprising the protozoan infrakingdoms Rhizaria [phyla Cercozoa and Re- taria (Radiozoa, Foraminifera)] and Excavata (phyla Loukozoa, Metamonada, Euglenozoa, Percolozoa), plus the kingdom Plantae [Viridaeplantae, Rhodophyta (sisters); Glaucophyta], the chromalveolate clade, and the protozoan phylum Apusozoa (Thecomonadea, Diphylleida). Chromalveolates comprise kingdom Chromista (Cryptista, Heterokonta, Haptophyta) and the protozoan infrakingdom Alveolata [phyla Cilio- phora and Miozoa (= Protalveolata, Dinozoa, Apicomplexa)], which diverged from a common ancestor that enslaved a red alga and evolved novel plastid protein-targeting machinery via the host rough ER and the enslaved algal plasma membrane (periplastid membrane). -

Entamoeba Invadens
R O UNDTAB LI Entamoeba invadens Entamoeba invadens is a very significant protozoan pathogen affecting several reptile taxons. Amoebiasis is often associated with disease in squamates, but can also cause significant morbidity and mortality in chelonians as well. This panel has extensive experience in chelonian medicine and will provide up-to-date information on diagnosing and treating chelonian species with amoebiasis. Barbara Bonner, DVM, MS The Turtle Hospital of New England 1 Grafton Road, Upton, MA 01568-1569, USA Tufts University School of Veterinary Medicine, North Grafton, MA 01536, USA Downloaded from http://meridian.allenpress.com/jhms/article-pdf/11/3/17/2203726/1529-9651_11_3_17.pdf by guest on 29 September 2021 Mary Denver, DVM Baltimore Zoo Druid Hill Park, Baltimore, MD 21217, USA Michael Gamer, DVM, DACVP Northwest Zoo Path 18210 Waverly, Snohomish, WA 98296, USA Charles Innis, VMD VC A Westboro Animal Hospital 155 Turnpike Road, Route 9, Westboro, MA 01581, USA Moderator: Robert Nathan, DVM 1). Which species of chelonians do you see with Entamoeba Geochelone elegans. We have seen clinical disease in mata invadens? matas, Chelus fimbriatus, and African mud turtles, Pelusios Bonner: I have seen Entamoeba and clinical signs of ill subniger. health that improved upon treatment in Gulf coast box turtle, Garner: Northwest ZooPath has cases of amoebiasis in all Terrapene Carolina major, three-toed box turtle, T. Carolina groups of reptiles, including snakes, lizards, chelonians, and triungulis, leopard tortoise, Geochelone pardalis, Travancore crocodilians. Since inception in 1994, we have accumulated tortoise, Indotestudo forsteni, Geoemyda yuwonoi, spiny tur 13 cases of amoebiasis in tortoises, and one case in a turtle. -

Amoebic Colitis Presenting As Subacute Intestinal Obstruction with Perforation
International Journal of TROPICAL DISEASE & Health 41(16): 58-62, 2020; Article no.IJTDH.61831 ISSN: 2278–1005, NLM ID: 101632866 Amoebic Colitis Presenting as Subacute Intestinal Obstruction with Perforation Renuka Verma1, Archana Budhwar1*, Priyanka Rawat1, Niti Dalal1, Anjali Bishlay1 and Sunita Singh1 1Pt. B. D. Sharma Post Graduate Institute of Medical Sciences, Rohtak-124001, Haryana, India. Authors’ contributions This work was carried out in collaboration among all authors. Authors RV and SS designed the study, reviewed the manuscript and edited it. Author Archana Budhwar wrote the protocol and wrote the first draft of the manuscript. Author PR managed the literature searches. Authors ND and Anjali Bishlay managed the analyses of the study. All authors read and approved the final manuscript. Article Information DOI: 10.9734/IJTDH/2020/v41i1630367 Editor(s): (1) Dr. Giuseppe Murdaca, University of Genoa, Italy. Reviewers: (1) Ammar M. Al-Aalim, Mosul University, Iraq. (2) Tarunbir Singh, Guru Angad Dev Veterinary & Animal Sciences University, Ludhiana. Complete Peer review History: http://www.sdiarticle4.com/review-history/61831 Received 10 August 2020 Case Study Accepted 16 October 2020 Published 12 November 2020 ABSTRACT Infestation with Entamoeba histolytica is worldwide, especially in developing areas. Presented case study included amoebic colitis in a 45 years old man complaining of abdominal distension and non- passage of stools since three days. Abdominal region was diffusely distended and tender in right iliac fossa. Plain abdominal radiography revealed prominent gut loops and minimal intergut free fluid. At laparotomy, malrotation of gut was present. Histopathological examination of intestinal samples confirmed final diagnosis of amoebic colitis post-operatively. -

Entamoeba Histolytica—Gut Microbiota Interaction: More Than Meets the Eye
microorganisms Review Entamoeba histolytica—Gut Microbiota Interaction: More Than Meets the Eye Serge Ankri Department of Molecular Microbiology, Ruth and Bruce Rappaport Faculty of Medicine, Haifa 31096, Israel; [email protected] Abstract: Amebiasis is a disease caused by the unicellular parasite Entamoeba histolytica. In most cases, the infection is asymptomatic but when symptomatic, the infection can cause dysentery and invasive extraintestinal complications. In the gut, E. histolytica feeds on bacteria. Increasing evidences support the role of the gut microbiota in the development of the disease. In this review we will discuss the consequences of E. histolytica infection on the gut microbiota. We will also discuss new evidences about the role of gut microbiota in regulating the resistance of the parasite to oxidative stress and its virulence. Keywords: gut microbiota; entamoeba histolytica; resistance to oxidative stress; resistance to nitrosative stress; virulence 1. Introduction Amebiasis is caused by the protozoan parasite Entamoeba histolytica. This disease is a significant hazard in underdeveloped countries with reduced socioeconomic and poor Citation: Ankri, S. Entamoeba sanitation. It is assessed that amebiasis accounted for 55,500 deaths and 2.237 million histolytica—Gut Microbiota disability-adjusted life years (the sum of years of life lost and years lived with disability) Interaction: More Than Meets the Eye. in 2010 [1]. Amebiasis has also been diagnosed in tourists from developed countries who Microorganisms 2021, 9, 581. return from vacation in endemic regions. Inflammation of the large intestine and liver https://doi.org/10.3390/ abscess represent the main clinical manifestations of amebiasis. Amebiasis is caused by the microorganisms9030581 ingestion of food contaminated with cysts, the infective form of the parasite.