Comparative Cell Biology in Diplomonads
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Introgression and Hybridization in Animal Parasites
Genes 2010, 1, 102-123; doi:10.3390/genes1010102 OPEN ACCESS genes ISSN 2073-4425 www.mdpi.com/journal/genes Review An Infectious Topic in Reticulate Evolution: Introgression and Hybridization in Animal Parasites Jillian T. Detwiler * and Charles D. Criscione Department of Biology, Texas A&M University, 3258 TAMU, College Station, TX 77843, USA; E-Mail: [email protected] * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +1-979-845-0925; Fax: +1-979-845-2891. Received: 29 April 2010; in revised form: 7 June 2010 / Accepted: 7 June 2010 / Published: 9 June 2010 Abstract: Little attention has been given to the role that introgression and hybridization have played in the evolution of parasites. Most studies are host-centric and ask if the hybrid of a free-living species is more or less susceptible to parasite infection. Here we focus on what is known about how introgression and hybridization have influenced the evolution of protozoan and helminth parasites of animals. There are reports of genome or gene introgression from distantly related taxa into apicomplexans and filarial nematodes. Most common are genetic based reports of potential hybridization among congeneric taxa, but in several cases, more work is needed to definitively conclude current hybridization. In the medically important Trypanosoma it is clear that some clonal lineages are the product of past hybridization events. Similarly, strong evidence exists for current hybridization in human helminths such as Schistosoma and Ascaris. There remain topics that warrant further examination such as the potential hybrid origin of polyploid platyhelminths. -

Pronunciation Guide to Microorganisms
Pronunciation Guide to Microorganisms This pronunciation guide is provided to aid each student in acquiring a greater ease in discussing, describing, and using specific microorganisms. Please note that genus and species names are italicized. If they cannot be italicized, then they should be underlined (example: a lab notebook). Prokaryotic Species Correct Pronunciation Acetobacter aceti a-se-toh-BAK-ter a-SET-i Acetobacter pasteurianus a-se-toh-BAK-ter PAS-ter-iann-us Acintobacter calcoacetius a-sin-ee-toe-BAK-ter kal-koh-a-SEE-tee-kus Aerococcus viridans (air-o)-KOK-kus vi-ree-DANS Agrobacterium tumefaciens ag-roh-bak-TEAR-ium too-me-FAY-she-ens Alcaligenes denitrificans al-KAHL-li-jen-eez dee-ni-TREE-fee-cans Alcaligenes faecalis al-KAHL-li-jen-eez fee-KAL-is Anabaena an-na-BEE-na Azotobacter vinelandii a-zoe-toe-BAK-ter vin-lan-DEE-i Bacillus anthracis bah-SIL-lus AN-thray-sis Bacillus lactosporus bah-SIL-lus LAK-toe-spore-us Bacillus megaterium bah-SIL-lus Meg-a-TEER-ee-um Bacillus subtilis bah-SIL-lus SA-til-us Borrelia recurrentis bore-RELL-ee-a re-kur-EN-tis Branhamella catarrhalis bran-hem-EL-ah cat-arr-RAH-lis Citrobacter freundii sit-roe-BACK-ter FROND-ee-i Clostridium perfringens klos-TREH-dee-um per-FRINGE-enz Clostridium sporogenes klos-TREH-dee-um spore-AH-gen-ease Clostridium tetani klos-TREH-dee-um TET-ann-ee Corynebacterium diphtheriae koh-RYNE-nee-back-teer-ee-um dif-THEE-ry-ee Corynebacterium hofmanni koh-RYNE-nee-back-teer-ee-um hoff-MAN-eye Corynebacterium xerosis koh-RYNE-nee-back-teer-ee-um zer-OH-sis Enterobacter -

Sex Is a Ubiquitous, Ancient, and Inherent Attribute of Eukaryotic Life
PAPER Sex is a ubiquitous, ancient, and inherent attribute of COLLOQUIUM eukaryotic life Dave Speijera,1, Julius Lukešb,c, and Marek Eliášd,1 aDepartment of Medical Biochemistry, Academic Medical Center, University of Amsterdam, 1105 AZ, Amsterdam, The Netherlands; bInstitute of Parasitology, Biology Centre, Czech Academy of Sciences, and Faculty of Sciences, University of South Bohemia, 370 05 Ceské Budejovice, Czech Republic; cCanadian Institute for Advanced Research, Toronto, ON, Canada M5G 1Z8; and dDepartment of Biology and Ecology, University of Ostrava, 710 00 Ostrava, Czech Republic Edited by John C. Avise, University of California, Irvine, CA, and approved April 8, 2015 (received for review February 14, 2015) Sexual reproduction and clonality in eukaryotes are mostly Sex in Eukaryotic Microorganisms: More Voyeurs Needed seen as exclusive, the latter being rather exceptional. This view Whereas absence of sex is considered as something scandalous for might be biased by focusing almost exclusively on metazoans. a zoologist, scientists studying protists, which represent the ma- We analyze and discuss reproduction in the context of extant jority of extant eukaryotic diversity (2), are much more ready to eukaryotic diversity, paying special attention to protists. We accept that a particular eukaryotic group has not shown any evi- present results of phylogenetically extended searches for ho- dence of sexual processes. Although sex is very well documented mologs of two proteins functioning in cell and nuclear fusion, in many protist groups, and members of some taxa, such as ciliates respectively (HAP2 and GEX1), providing indirect evidence for (Alveolata), diatoms (Stramenopiles), or green algae (Chlor- these processes in several eukaryotic lineages where sex has oplastida), even serve as models to study various aspects of sex- – not been observed yet. -

3.2.2 Diplomonad (Hexamitid) Flagellates - 1
3.2.2 Diplomonad (Hexamitid) Flagellates - 1 3.2.2 Diplomonad (Hexamitid) Flagellates: Diplomonadiasis, Hexamitosis, Spironucleosis Sarah L. Poynton Department of Comparative Medicine Johns Hopkins University School of Medicine 1-127 Jefferson Building, Johns Hopkins Hospital 600 North Wolfe Street Baltimore, MD 21287 410/502-5065 fax: 443/287-2954 [email protected] [email protected] A. Name of Disease and Etiological Agent Diplomonadiasis or hexamitosis is infection by diplomonad flagellates (Order Diplomonadida, suborder Diplomonadina, Family Hexamitidae). If the exact genus is known, the infections may be reported as hexamitiasis (Hexamita), octomitosis (Octomitus), or spironucleosis (Spironucleus); of these, probably only the latter is applicable to fish (see below). Infections may be reported as localized (commonly in the intestine, and possibly also including “hole-in-the head disease” of cichlids (Paull and Matthews 2001), or disseminated or systemic (Ferguson and Moccia 1980; Kent et al. 1992; Poppe et al. 1992; Sterud et al. 1998). Light microscopy studies have reported three genera from fish - namely Hexamita, Octomitus, and Spironucleus. However, transmission electron microscopy (TEM) is needed to confirm genus (Poynton and Sterud 2002), and light microscopy studies are therefore taxonomically unreliable. If TEM is not available, the organisms should be recorded as diplomonad or hexamitid flagellates. All recent comprehensive ultrastructural studies show only the genus Spironucleus infecting fish, and it is probable that this is the genus to which all diplomonads from fish belong (Poynton and Sterud 2002). Some 15 to 20 species of diplomonads have been reported from fish (Poynton and Sterud 2002). However, most descriptions do not include comprehensive surface and internal ultrastructure and thus are incomplete. -

Cyprinus Carpio
Cyprinus carpio Investigation of infection by some Endo- parasitic Protozoa species in common carp ( Cyprinus carpio ) in AL-Sinn fishfarm 2014 Cyprinus carpio Investigation of infection by some Endo- parasitic Protozoa species in common carp ( Cyprinus carpio ) in AL-Sinn fishfarm 2014 I IV V VI VII 2 1 3 2 3 3 4 4 4 1 4 4 1 1 4 2 4 6 6 1 2 4 6 6 7 8 8 10 12 14 15 15 16 2 2 4 I 16 16 16 17 17 18 18 19 19 19 Hexamitosis 3 2 4 19 19 20 20 21 21 21 1 3 4 22 23 2 3 4 26 5 26 1 5 27 2 5 28 3 5 29 4 5 29 1 4 5 29 2 4 5 33 3 4 5 II 34 4 4 5 34 5 4 5 36 6 36 1 6 42 2 6 42 Hexamitosis 3 6 43 7 44 8 44 9 49 10 49 1 10 51 2 10 III 5 1 Oocyst 7 2 15 3 11 Merozoites 4 12 5 12 6 13 7 14 8 15 9 Trypanosoma 1 18 10 Trypanoplasma 2 19 11 21 Hexamita 12 23 Hexamita 13 A 28 14 B 30 Cyprinus carpio carpio 15 31 16 33 17 34 18 34 19 35 20 35 21 36 22 IV 40 Goussia carpelli 23 40 Macrogametocyte 24 41 Merozoites 25 37 1 Goussia carpelli 39 2 Goussia 41 3 carpelli Goussia 42 carpelli 4 43 5 Goussia 44 carpelli 6 V Goussia Trypanosoma Trypanoplasma borreli Goussia subepithelialis carpelli Hexamita intestinalis danilewskyi Hexamitosis 200 2013 / 5 / 14 2012 / 6 / 6 Goussia carpelli Goussia subepithelials Nodular Coccidiosis Macrogametocytes Merozoites 3 Goussia carpelli 5.4 1.6 17.6 95.84 17.5 16 Trypanosoma Trypanoplasma borreli Hexamita intestinalis danilewskyi VI Abstract This study aimed at investigating the infection of cultured common carp ( Cyprinus carpio L. -

Multigene Eukaryote Phylogeny Reveals the Likely Protozoan Ancestors of Opis- Thokonts (Animals, Fungi, Choanozoans) and Amoebozoa
Accepted Manuscript Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opis- thokonts (animals, fungi, choanozoans) and Amoebozoa Thomas Cavalier-Smith, Ema E. Chao, Elizabeth A. Snell, Cédric Berney, Anna Maria Fiore-Donno, Rhodri Lewis PII: S1055-7903(14)00279-6 DOI: http://dx.doi.org/10.1016/j.ympev.2014.08.012 Reference: YMPEV 4996 To appear in: Molecular Phylogenetics and Evolution Received Date: 24 January 2014 Revised Date: 2 August 2014 Accepted Date: 11 August 2014 Please cite this article as: Cavalier-Smith, T., Chao, E.E., Snell, E.A., Berney, C., Fiore-Donno, A.M., Lewis, R., Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa, Molecular Phylogenetics and Evolution (2014), doi: http://dx.doi.org/10.1016/ j.ympev.2014.08.012 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. 1 1 Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts 2 (animals, fungi, choanozoans) and Amoebozoa 3 4 Thomas Cavalier-Smith1, Ema E. Chao1, Elizabeth A. Snell1, Cédric Berney1,2, Anna Maria 5 Fiore-Donno1,3, and Rhodri Lewis1 6 7 1Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK. -

A Wide Diversity of Previously Undetected Freeliving
Environmental Microbiology (2010) 12(10), 2700–2710 doi:10.1111/j.1462-2920.2010.02239.x A wide diversity of previously undetected free-living relatives of diplomonads isolated from marine/saline habitatsemi_2239 2700..2710 Martin Kolisko,1 Jeffrey D. Silberman,2 Kipferlia n. gen. The remaining isolates include rep- Ivan Cepicka,3 Naoji Yubuki,4† Kiyotaka Takishita,5 resentatives of three other lineages that likely repre- Akinori Yabuki,4 Brian S. Leander,6 Isao Inouye,4 sent additional undescribed genera (at least). Small- Yuji Inagaki,7 Andrew J. Roger8 and subunit ribosomal RNA gene phylogenies show that Alastair G. B. Simpson1* CLOs form a cloud of six major clades basal to the Departments of 1Biology and 8Biochemistry and diplomonad-retortamonad grouping (i.e. each of the Molecular Biology, Dalhousie University, Halifax, Nova six CLO clades is potentially as phylogenetically Scotia, Canada. distinct as diplomonads and retortamonads). CLOs 2Department of Biological Sciences, University of will be valuable for tracing the evolution of Arkansas, Fayetteville, AR, USA. diplomonad cellular features, for example, their 3Department of Zoology, Faculty of Science, Charles extremely reduced mitochondrial organelles. It is University in Prague, Prague, Czech Republic. striking that the majority of CLO diversity was unde- 4Institute of Biological Sciences, Graduate School of Life tected by previous light microscopy surveys and and Environmental Sciences and 7Center for environmental PCR studies, even though they inhabit Computational Sciences and Institute of Biological a commonly sampled environment. There is no Sciences, University of Tsukuba, Tsukuba, Ibaraki, reason to assume this is a unique situation – it is Japan. likely that undersampling at the level of major lin- 5Japan Agency for Marine-Earth Science and eages is still widespread for protists. -
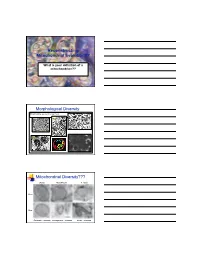
Reconstructing Mitochondrial Evolution?? Morphological Diversity Mitochondrial Diversity???
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial Diversity??? Chicken Neocallimastix T. foetus 100 nm 50 nm Entamoeba - mitosome microsporidian - mitosome Giardia - mitosome Reconstructing Evolution Mitochondrial evolution well established endosymbiotic theory α-proteobacterium - Rickettsia prowazekii Hydrogenosomal evolution No DNA NOW 2 examples Nyctotherus and Blastocystis (MLO) Several proteins similar to mitochondria Mitosome evolution No DNA Few proteins identified similar to mitochondria Origins via Endosymbiosis Current dogma - mitochondria and related Aerobic α-proteobacterium prokaryote gave rise to present day mitochondria. organelles arose just once in evolution Are hydrogenosomes and mitosomes of anaerobic protists derived from the same proto-mitochondrion? Evidence for: accumulating evidence for several proteins that are currently found in mitochondria - Proteins of Fe-S cluster formation. Scenario A Common ancestor facultative Scenario B organism? degenerate mitochondrion invoke lateral gene transfer from anaerobic prokaryotes Hypotheses for Mito Acquisition Mitochondrion-related Organelles 2 way to interpret this summary - can you think of both? Groups with mitochondrial homologues Hjort et al Phil. Trans. R. Soc. B 2010 365, 713-727 Origin of Hydrogenosomes (Mitosomes?) 1. a)Conversion of Mitochondria HYD b)Common ancestor with mitochondria ? HYD 2. Independent Origin α-proteobacterium ? - anaerobic bacterium HYD Organelles - origins and biogenesis Approaches: (1) Conduct phylogenetic analyses of similar proteins Hsp70 Fd Hsp60 Isc subunits (2) Examine protein targeting to the organelle matrix protein targeting membrane protein targeting (3) Characterize membrane/translocation components These components could have evolved as the endosymbiont was converted to organelle. Reveals evolutionary history. -

Superorganisms of the Protist Kingdom: a New Level of Biological Organization
Foundations of Science https://doi.org/10.1007/s10699-020-09688-8 Superorganisms of the Protist Kingdom: A New Level of Biological Organization Łukasz Lamża1 © The Author(s) 2020 Abstract The concept of superorganism has a mixed reputation in biology—for some it is a conveni- ent way of discussing supra-organismal levels of organization, and for others, little more than a poetic metaphor. Here, I show that a considerable step forward in the understand- ing of superorganisms results from a thorough review of the supra-organismal levels of organization now known to exist among the “unicellular” protists. Limiting the discussion to protists has enormous advantages: their bodies are very well studied and relatively sim- ple (as compared to humans or termites, two standard examples in most discussions about superorganisms), and they exhibit an enormous diversity of anatomies and lifestyles. This allows for unprecedented resolution in describing forms of supra-organismal organiza- tion. Here, four criteria are used to diferentiate loose, incidental associations of hosts with their microbiota from “actual” superorganisms: (1) obligatory character, (2) specifc spatial localization of microbiota, (3) presence of attachment structures and (4) signs of co-evolu- tion in phylogenetic analyses. Three groups—that have never before been described in the philosophical literature—merit special attention: Symbiontida (also called Postgaardea), Oxymonadida and Parabasalia. Specifcally, it is argued that in certain cases—for Bihos- pites bacati and Calkinsia aureus (symbiontids), Streblomastix strix (an oxymonad), Joe- nia annectens and Mixotricha paradoxa (parabasalids) and Kentrophoros (a ciliate)—it is fully appropriate to describe the whole protist-microbiota assocation as a single organism (“superorganism”) and its elements as “tissues” or, arguably, even “organs”. -

Protist Phylogeny and the High-Level Classification of Protozoa
Europ. J. Protistol. 39, 338–348 (2003) © Urban & Fischer Verlag http://www.urbanfischer.de/journals/ejp Protist phylogeny and the high-level classification of Protozoa Thomas Cavalier-Smith Department of Zoology, University of Oxford, South Parks Road, Oxford, OX1 3PS, UK; E-mail: [email protected] Received 1 September 2003; 29 September 2003. Accepted: 29 September 2003 Protist large-scale phylogeny is briefly reviewed and a revised higher classification of the kingdom Pro- tozoa into 11 phyla presented. Complementary gene fusions reveal a fundamental bifurcation among eu- karyotes between two major clades: the ancestrally uniciliate (often unicentriolar) unikonts and the an- cestrally biciliate bikonts, which undergo ciliary transformation by converting a younger anterior cilium into a dissimilar older posterior cilium. Unikonts comprise the ancestrally unikont protozoan phylum Amoebozoa and the opisthokonts (kingdom Animalia, phylum Choanozoa, their sisters or ancestors; and kingdom Fungi). They share a derived triple-gene fusion, absent from bikonts. Bikonts contrastingly share a derived gene fusion between dihydrofolate reductase and thymidylate synthase and include plants and all other protists, comprising the protozoan infrakingdoms Rhizaria [phyla Cercozoa and Re- taria (Radiozoa, Foraminifera)] and Excavata (phyla Loukozoa, Metamonada, Euglenozoa, Percolozoa), plus the kingdom Plantae [Viridaeplantae, Rhodophyta (sisters); Glaucophyta], the chromalveolate clade, and the protozoan phylum Apusozoa (Thecomonadea, Diphylleida). Chromalveolates comprise kingdom Chromista (Cryptista, Heterokonta, Haptophyta) and the protozoan infrakingdom Alveolata [phyla Cilio- phora and Miozoa (= Protalveolata, Dinozoa, Apicomplexa)], which diverged from a common ancestor that enslaved a red alga and evolved novel plastid protein-targeting machinery via the host rough ER and the enslaved algal plasma membrane (periplastid membrane). -

Identification of the Meiotic Life Cycle Stage of Trypanosoma Brucei in The
Identification of the meiotic life cycle stage of Trypanosoma brucei in the tsetse fly Lori Peacocka,b, Vanessa Ferrisa,b, Reuben Sharmac,1, Jack Sunterc, Mick Baileyb, Mark Carringtonc, and Wendy Gibsona,2 aSchool of Biological Sciences, University of Bristol, Bristol BS8 1UG, United Kingdom; bDepartment of Clinical Veterinary Science, University of Bristol, Bristol BS40 7DU, United Kingdom; and cDepartment of Biochemistry, University of Cambridge, Cambridge CB2 1QW, United Kingdom Edited by Francisco J. Ayala, University of California, Irvine, CA, and approved January 27, 2011 (received for review December 23, 2010) Elucidating the mechanism of genetic exchange is fundamental for genetic exchange in T. brucei involves mixing of mitochondrial understanding how genes for such traits as virulence, disease (kinetoplast) and nuclear genomes, because hybrid progeny have phenotype, and drug resistance are transferred between pathogen hybrid kinetoplast DNA (kDNA) networks with mini-circles de- strains. Genetic exchange occurs in the parasitic protists Trypano- rived from both parents (14, 15). Plausible models for the gen- soma brucei, T. cruzi, and Leishmania major, but the precise cellular eration of hybrid kDNA networks are limited by the complex mechanisms are unknown, because the process has not been ob- structure and highly ordered replication of this concatenated served directly. Here we exploit the identification of homologs of mass of small DNA circles (16). Finally, trypanosomes belong to meiotic genes in the T. brucei genome and demonstrate that three the Euglenozoa, a deep branch within the excavate eukaryote su- functionally distinct, meiosis-specific proteins are expressed in the pergroup (17, 18). The production of four haploid gametes and nucleus of a single specific cell type, defining a previously unde- subsequent fusion to reform the diploid occurring in trypanosomes scribed developmental stage occurring within the tsetse flysalivary would strongly suggest the presence of a typical meiosis in the last gland. -

Molecular Identification and Evolution of Protozoa Belonging to the Parabasalia Group and the Genus Blastocystis
UNIVERSITAR DEGLI STUDI DI SASSARI SCUOLA DI DOTTORATO IN SCIENZE BIOMOLECOLARI E BIOTECNOLOGICHE (Intenational PhD School in Biomolecular and Biotechnological Sciences) Indirizzo: Microbiologia molecolare e clinica Molecular identification and evolution of protozoa belonging to the Parabasalia group and the genus Blastocystis Direttore della scuola: Prof. Masala Bruno Relatore: Prof. Pier Luigi Fiori Correlatore: Dott. Eric Viscogliosi Tesi di Dottorato : Dionigia Meloni XXIV CICLO Nome e cognome: Dionigia Meloni Titolo della tesi : Molecular identification and evolution of protozoa belonging to the Parabasalia group and the genus Blastocystis Tesi di dottorato in scienze Biomolecolari e biotecnologiche. Indirizzo: Microbiologia molecolare e clinica Universit degli studi di Sassari UNIVERSITAR DEGLI STUDI DI SASSARI SCUOLA DI DOTTORATO IN SCIENZE BIOMOLECOLARI E BIOTECNOLOGICHE (Intenational PhD School in Biomolecular and Biotechnological Sciences) Indirizzo: Microbiologia molecolare e clinica Molecular identification and evolution of protozoa belonging to the Parabasalia group and the genus Blastocystis Direttore della scuola: Prof. Masala Bruno Relatore: Prof. Pier Luigi Fiori Correlatore: Dott. Eric Viscogliosi Tesi di Dottorato : Dionigia Meloni XXIV CICLO Nome e cognome: Dionigia Meloni Titolo della tesi : Molecular identification and evolution of protozoa belonging to the Parabasalia group and the genus Blastocystis Tesi di dottorato in scienze Biomolecolari e biotecnologiche. Indirizzo: Microbiologia molecolare e clinica Universit degli studi di Sassari Abstract My thesis was conducted on the study of two groups of protozoa: the Parabasalia and Blastocystis . The first part of my work was focused on the identification, pathogenicity, and phylogeny of parabasalids. We showed that Pentatrichomonas hominis is a possible zoonotic species with a significant potential of transmission by the waterborne route and could be the aetiological agent of gastrointestinal troubles in children.