202 Common Redshank Put Your Logo Here
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nordmann's Greenshank Population Analysis, at Pantai Cemara Jambi
Final Report Nordmann’s Greenshank Population Analysis, at Pantai Cemara Jambi Cipto Dwi Handono1, Ragil Siti Rihadini1, Iwan Febrianto1 and Ahmad Zulfikar Abdullah1 1Yayasan Ekologi Satwa Alam Liar Indonesia (Yayasan EKSAI/EKSAI Foundation) Surabaya, Indonesia Background Many shorebirds species have declined along East Asian-Australasian Flyway which support the highest diversity of shorebirds in the world, including the globally endangered species, Nordmann’s Greenshank. Nordmann’s Greenshank listed as endangered in the IUCN Red list of Threatened Species because of its small and declining population (BirdLife International, 2016). It’s one of the world’s most threatened shorebirds, is confined to the East Asian–Australasian Flyway (Bamford et al. 2008, BirdLife International 2001, 2012). Its global population is estimated at 500–1,000, with an estimated 100 in Malaysia, 100–200 in Thailand, 100 in Myanmar, plus unknown but low numbers in NE India, Bangladesh and Sumatra (Wetlands International 2006). The population is suspected to be rapidly decreasing due to coastal wetland development throughout Asia for industry, infrastructure and aquaculture, and the degradation of its breeding habitat in Russia by grazing Reindeer Rangifer tarandus (BirdLife International 2012). Mostly Nordmann’s Greenshanks have been recorded in very small numbers throughout Southeast Asia, and there are few places where it has been reported regularly. In Myanmar, for example, it was rediscovered after a gap of almost 129 years. The total count recorded by the Asian Waterbird Census (AWC) in 2006 for Myanmar was 28 birds with 14 being the largest number at a single locality (Naing 2007). In 2011–2012, Nordmann’s Greenshank was found three times in Sumatera Utara province, N Sumatra. -

Population Analysis and Community Workshop for Far Eastern Curlew Conservation Action in Pantai Cemara, Desa Sungai Cemara – Jambi
POPULATION ANALYSIS AND COMMUNITY WORKSHOP FOR FAR EASTERN CURLEW CONSERVATION ACTION IN PANTAI CEMARA, DESA SUNGAI CEMARA – JAMBI Final Report Small Grant Fund of the EAAFP Far Eastern Curlew Task Force Iwan Febrianto, Cipto Dwi Handono & Ragil S. Rihadini Jambi, Indonesia 2019 The aim of this project are to Identify the condition of Far Eastern Curlew Population and the remaining potential sites for Far Eastern Curlew stopover in Sumatera, Indonesia and protect the remaining stopover sites for Far Eastern Curlew by educating the government, local people and community around the sites as the effort of reducing the threat of habitat degradation, habitat loss and human disturbance at stopover area. INTRODUCTION The Far Eastern Curlew (Numenius madagascariencis) is the largest shorebird in the world and is endemic to East Asian – Australian Flyway. It is one of the Endangered migratory shorebird with estimated global population at 38.000 individual, although a more recent update now estimates the population at 32.000 (Wetland International, 2015 in BirdLife International, 2017). An analysis of monitoring data collected from around Australia and New Zealand (Studds et al. in prep. In BirdLife International, 2017) suggests that the species has declined much more rapidly than was previously thought; with an annual rate of decline of 0.058 equating to a loss of 81.7% over three generations. Habitat loss occuring as a result of development is the most significant threat currently affecting migratory shorebird along the EAAF (Melville et al. 2016 in EAAFP 2017). Loss of habitat at critical stopover sites in the Yellow Sea is suspected to be the key threat to this species and given that it is restricted to East Asian - Australasian Flyway, the declines in the non-breeding are to be representative of the global population. -

Trends in Breeding Phenology Across Ten Decades Show Varying Adjustments to Environmental Changes in Four Wader Species
Bird Study ISSN: 0006-3657 (Print) 1944-6705 (Online) Journal homepage: http://www.tandfonline.com/loi/tbis20 Trends in breeding phenology across ten decades show varying adjustments to environmental changes in four wader species Hans Meltofte, Ole Amstrup, Troels Leuenhagen Petersen, Frank Rigét & Anders P. Tøttrup To cite this article: Hans Meltofte, Ole Amstrup, Troels Leuenhagen Petersen, Frank Rigét & Anders P. Tøttrup (2018) Trends in breeding phenology across ten decades show varying adjustments to environmental changes in four wader species, Bird Study, 65:1, 44-51, DOI: 10.1080/00063657.2018.1444014 To link to this article: https://doi.org/10.1080/00063657.2018.1444014 Published online: 20 Mar 2018. Submit your article to this journal Article views: 49 View related articles View Crossmark data Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=tbis20 BIRD STUDY, 2018 VOL. 65, NO. 1, 44–51 https://doi.org/10.1080/00063657.2018.1444014 Trends in breeding phenology across ten decades show varying adjustments to environmental changes in four wader species Hans Meltoftea, Ole Amstrupb, Troels Leuenhagen Petersenc, Frank Rigéta and Anders P. Tøttrupc aDepartment of Bioscience, Aarhus University, Roskilde, Denmark; bAmphi Consult, Aalborg, Denmark; cCenter for Macroecology, Evolution and Climate, University of Copenhagen, Copenhagen, Denmark ABSTRACT ARTICLE HISTORY Capsule: During 1928–2016, initiation of egg-laying advanced in two wader species, remained Received 8 May 2017 unchanged in one, and was delayed in one species. The changes across years and variation Accepted 19 February 2018 among species can be explained by climatic variables and differences in migratory strategies. -

Tringarefs V1.3.Pdf
Introduction I have endeavoured to keep typos, errors, omissions etc in this list to a minimum, however when you find more I would be grateful if you could mail the details during 2016 & 2017 to: [email protected]. Please note that this and other Reference Lists I have compiled are not exhaustive and best employed in conjunction with other reference sources. Grateful thanks to Graham Clarke (http://grahamsphoto.blogspot.com/) and Tom Shevlin (www.wildlifesnaps.com) for the cover images. All images © the photographers. Joe Hobbs Index The general order of species follows the International Ornithologists' Union World Bird List (Gill, F. & Donsker, D. (eds). 2016. IOC World Bird List. Available from: http://www.worldbirdnames.org/ [version 6.1 accessed February 2016]). Version Version 1.3 (March 2016). Cover Main image: Spotted Redshank. Albufera, Mallorca. 13th April 2011. Picture by Graham Clarke. Vignette: Solitary Sandpiper. Central Bog, Cape Clear Island, Co. Cork, Ireland. 29th August 2008. Picture by Tom Shevlin. Species Page No. Greater Yellowlegs [Tringa melanoleuca] 14 Green Sandpiper [Tringa ochropus] 16 Greenshank [Tringa nebularia] 11 Grey-tailed Tattler [Tringa brevipes] 20 Lesser Yellowlegs [Tringa flavipes] 15 Marsh Sandpiper [Tringa stagnatilis] 10 Nordmann's Greenshank [Tringa guttifer] 13 Redshank [Tringa totanus] 7 Solitary Sandpiper [Tringa solitaria] 17 Spotted Redshank [Tringa erythropus] 5 Wandering Tattler [Tringa incana] 21 Willet [Tringa semipalmata] 22 Wood Sandpiper [Tringa glareola] 18 1 Relevant Publications Bahr, N. 2011. The Bird Species / Die Vogelarten: systematics of the bird species and subspecies of the world. Volume 1: Charadriiformes. Media Nutur, Minden. Balmer, D. et al 2013. Bird Atlas 2001-11: The breeding and wintering birds of Britain and Ireland. -

Iucn Red Data List Information on Species Listed On, and Covered by Cms Appendices
UNEP/CMS/ScC-SC4/Doc.8/Rev.1/Annex 1 ANNEX 1 IUCN RED DATA LIST INFORMATION ON SPECIES LISTED ON, AND COVERED BY CMS APPENDICES Content General Information ................................................................................................................................................................................................................................ 2 Species in Appendix I ............................................................................................................................................................................................................................... 3 Mammalia ............................................................................................................................................................................................................................................ 4 Aves ...................................................................................................................................................................................................................................................... 7 Reptilia ............................................................................................................................................................................................................................................... 12 Pisces ................................................................................................................................................................................................................................................. -

Migration Timing, Routes, and Connectivity of Eurasian Woodcock Wintering in Britain and Ireland
Migration Timing, Routes, and Connectivity of Eurasian Woodcock Wintering in Britain and Ireland ANDREW N. HOODLESS,1 Game & Wildlife Conservation Trust, Burgate Manor, Fordingbridge, Hampshire SP6 1EF, UK CHRISTOPHER J. HEWARD, Game & Wildlife Conservation Trust, Burgate Manor, Fordingbridge, Hampshire SP6 1EF, UK ABSTRACT Migration represents a critical time in the annual cycle of Eurasian woodcock (Scolopax rusticola), with poten- tial consequences for individual fitness and survival. In October–December, Eurasian woodcock migrate from breeding grounds in northern Eurasia over thousands of kilometres to western Europe, returning in March–May. The species is widely hunted in Europe, with 2.3–3.5 million individuals shot per year; hence, an understanding of the timing of migra- tion and routes taken is an essential part of developing sustainable flyway management. Our aims were to determine the timing and migration routes of Eurasian woodcock wintering in Britain and Ireland, and to assess the degree of connec- tivity between breeding and wintering sites. We present data from 52 Eurasian woodcock fitted with satellite tags in late winter 2012–2016, which indicate that the timing of spring departure varied annually and was positively correlated with temperature, with a mean departure date of 26 March (± 1.4 days SE). Spring migration distances averaged 2,851 ± 165 km (SE), with individuals typically making 5 stopovers. The majority of our sample of tagged Eurasian woodcock migrated to breeding sites in northwestern Russia (54%), with smaller proportions breeding in Denmark, Scandinavia, and Finland (29%); Poland, Latvia, and Belarus (9.5%); and central Russia (7.5%). The accumulated migration routes of tagged individ- uals suggest a main flyway for Eurasian woodcock wintering in Britain and Ireland through Belgium, the Netherlands, and Germany, and then dividing to pass through the countries immediately north and south of the Baltic Sea. -
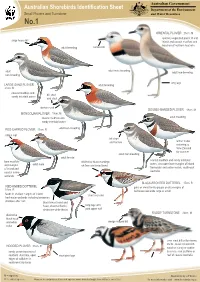
Australian Shorebirds Identification Sheet Department of the Environment Small Plovers and Turnstone and Water Resources No.1
Australian Government Australian Shorebirds Identification Sheet Department of the Environment Small Plovers and Turnstone and Water Resources No.1 ORIENTAL PLOVER 25cm. M sparsely vegetated plains of arid large heavy bill inland and coastal mudflats and beaches of northern Australia adult breeding narrow bill adult male breeding adult adult non-breeding non-breeding long legs LARGE SAND PLOVER adult breeding 21cm. M coastal mudflats and bill short sandy intertidal zones and stout darker mask DOUBLE-BANDED PLOVER 19cm. M MONGOLIAN PLOVER 19cm. M coastal mudflats and adult breeding sandy intertidal zones RED-CAPPED PLOVER 15cm. R adult non-breeding rufous cap bill short and narrow winter visitor returning to New Zealand for summer adult non-breeding adult female coastal mudflats and sandy intertidal bare mudflats distinctive black markings zones, also open bare margins of inland and margins adult male on face and breastband of inland and freshwater and saline marsh, south-east coastal saline Australia wetlands BLACK-FRONTED DOTTEREL 17cm. R RED-KNEED DOTTEREL pairs or small family groups on dry margins of 18cm. R feshwater wetlands large or small feeds in shallow margins of inland short rear end freshwater wetlands including temporary shallows after rain black breast band and head, chestnut flanks, long legs with distinctive white throat pink upper half RUDDY TURNSTONE 23cm. M distinctive black hood and white wedge shaped bill collar uses stout bill to flip stones, shells, seaweed and drift- 21cm. R HOODED PLOVER wood on sandy or cobble sandy ocean beaches of beaches, rock platform or southern Australia, open short pink legs reef of coastal Australia edges of saltlakes in south-west Australia M = migratory . -

Threatened Species List Spain
THREATENED SPECIES LIST SPAIN Threatened species included in the national inventory of the Ministry of MARM and/or in the Red List of the International Union for Conservation of Nature (IUCN) that are or may be inhabited in the areas of our Hydro Power Stations. 6 CRITIC ENDANGERED SPECIES (CR) GROUP SPECIE COMMON NAME CATEGORY (MARM) (IUCN) Birds Neophron percnopterus Egyptian Vulture CR EN Botaurus stellaris Great Bittern CR LC Mammals Lynx pardinus Iberian Lynx CR CR Ursus arctos Brown Bear CR (Northern Spain) LC Invertebrates Belgrandiella galaica Gastropoda CR No listed Macromia splendens Splendid Cruiser CR VU 24 ENDANGERED SPECIES (EN) GROUP SPECIE COMMON NAME CATEGORY (MARM) (IUCN) Amphibians Rana dalmatina Agile Frog EN LC Birds Pyrrhocorax pyrrhocorax Chough EN LC Hieraaetus fasciatus Bonelli´s Eagle EN LC Alectoris rufa Barbary Partridge EN LC Parus caeruleus Blue Tit EN LC Tyto alba Barn Owl EN LC Burhinus oedicnemus Stone Curlew EN LC Corvus corax Common Raven EN LC Chersophilus duponti Dupont´s Lark EN NT Milvus milvus Red Kite EN NT Aquila adalberti Spanish Imperial Eagle EN VU Cercotrichas galactotes Alzacola EN LC Reptiles Algyroides marchi Spanish Algyroides EN EN Emys orbicularis European Pond Turtle EN NT Mammals Rhinolophus mehelyi Mehely´s Horseshoe Bat EN VU Mustela lutreola European Mink EN EN Myotis capaccinii Long –Fingered bat EN VU Freshwater fish Salaria fluviatilis Freshwater blenny EN LC Chondrostoma turiense Madrija (Endemic) EN EN Cobitis vettonica Colmilleja del Alagón EN EN (Endemic) Invertebrates Gomphus -

Population Ecology of the Long-Billed Curlew (Numenius Americanus) in Western Idaho
POPULATION ECOLOGY OF THE LONG-BILLED CURLEW (NUMENIUS AMERICANUS) IN WESTERN IDAHO ROLAND L. REDMOND AND DONALD A. JENNI Departmentof Zoology,University of Montana,Missoula, Montana 59812 USA ABSTRACT.--IIlwestern Idaho, a breeding population of Long-billed Curlews (Numenius americanus)remained relatively stable over a 7-yr period.Productivity was monitored closely from 1977through 1979. Clutch mortality rates did not differ significantlyamong years, and meanclutch survival for a 32-daynesting period (4 daysegg laying and 28 daysincubation) was40% for all 3 yr combined.Females laid justone clutch(usually of 4 eggs)each season, and meanclutch size was significantlysmaller in 1979than in 1977and 1978.Estimated fledgingsuccess ranged from 0.40in 1977to 0.17in 1978and wasconsistently greater for femalesthat nestedearly eachseason. Mean annualadult survivalwas estimatedat 85% basedon resightingsof color-markedindividuals. Limited data for subadultsurvival pre- cludedcomplete demographic analysis. Nevertheless, given the estimatesof productivity and adult survivalpooled for 3 yr, survivalof subadultsfrom fledginguntil first breeding as3-year-olds needed to beonly 58% to maintainthe stablepopulation size observed through 1983. Received10 June1985, accepted 7 April 1986. THE population ecology of shorebirds (Cha- land et al. 1982), and Common Redshank (Yates radrii) is not well known, probably because 1982). Larger speciestend to be better studied many speciesare wary, breed in remote areas, during the nonbreeding season,and survival or both. Most thoroughly studied are species estimatesbased on annual returns to wintering that nest at temperate latitudes such as the Eur- grounds exist for Bar-tailed Godwits (Limosa asianOystercatcher (Haematopus ostralegus; Goss- lapponica)and Eurasian Curlews (Numeniusar- Custard et al. 1982, Safriel et al. 1984), Common quata) (Evans and Pienkowski 1984). -

Tringa Glareola 2000&Rdquo
30 Workshop on Project "Tringa glareola 2000" and Seminar "Research on Wader Migration in Poland"- 21-22 November 1998, Gdynia, Poland. Magdalena Remisiewicz& Mateusz Sciborski Remisiewicz,M. & Sciborski,M. 1999.Workshop on Project"Tringa glareola 2000" andSeminar "Research on WaderMigration m Poland"- 21-22 November 1998, Gdynia, Poland.Wader Study Group Bull. 89:30 - 31 MagdalenaRemisiewicz & MateuszSciborski, Waterbird Research Group "KULING", c/o Dept. of VertebrateZoology and Ecology,Univ. of Gdansk,al. Legion6w9, 80-441 Gdansk,Poland,' E-mail.' [email protected] The WaderStudy Groupprogramme (Poland)by the WRG "KULING". keen "KULING" bird-ringers "Tringaglareola 2000", co-ordinated During the meeting,participants from succeededin catchinga male Mallard, by the WaterbirdResearch Group Poland, Sweden, Romania, Estonia and triumphantlygiving it the name "KULING" has developedremarkably Ukrainepresented the resultsof pastand "Conference". Other attractions sinceits startin 1997. Its main purpose, presentstudies on the speciesin their providedfor the guestswere two to definethe migrationroutes of Wood countries. A common set of exhibitionsprepared especially for the Sandpiperand investigate its migration measurements to be taken from Wood workshop.The "Museumof WRG strategy,seems to be met by the Sandpiperswas agreed,including new KULING" presentedhistorical pieces of methodsused. They include:colour measures(primary wear, fat score)to be equipmentused during fieldwork or ringing,dyeing, counts and tested.Topic groups, expressing specimensreferring to important observations(for more detailed individualinterests in analysing momentsin the historyof the group descriptionof the project- seeWSG particularproblems; migration (someof the exhibitswere 15 years Bull. 84: 21-22). The network of sites dynamics,recoveries, biometrics, moult, old). The other was a collection of contributinghas been growing refuelhngstrategy, were established. -

Biometrics and Breeding Phenology of Terek Sandpipers in the Pripyat’ Valley, S Belarus
54 Wader Study Group Bulletin Biometrics and breeding phenology of Terek Sandpipers in the Pripyat’ Valley, S Belarus NATALIA KARLIONOVA1, MAGDALENA REMISIEWICZ2, PAVEL PINCHUK1 1Institute of Zoology, Belarussian National Academy of Sciences, Academichnaya Str. 27, 220072 Minsk, Belarus. [email protected] 2Avian Ecophysiology Unit, Dept of Vertebrate Ecology and Zoology, Univ. of Gdansk, al. Legionów 9, 80-441 Gdansk, Poland. [email protected] Karlionova, N., Remisiewicz, M. & Pinchuk, P. 2006. Biometrics and breeding phenology of Terek Sandpipers in the Pripyat’ Valley, S Belarus. Wader Study Group Bull. 110: 54–58. Keywords: Terek Sandpiper, Xenus cinereus, biometrics, breeding phenology, Pripyat’ Valley, Belarus We present data on the breeding phenology and biometrics of Terek Sandpipers from the isolated westernmost population in the Pripyat’ river valley, S Belarus, close to the border with Ukraine. Studies were conducted on floodplain islands between the beginning of April and mid-July during 1996–1999 and 2002–2006. Over the years, the first arrivals appeared during 10–26 April (median 14 April), first eggs were laid during 24 April to 5 May (median 30 April) and the latest egg was laid on 25 May, first chicks hatched during 19 May to 1 June (median 25 May) and the first fledged juveniles were caught on 23 June. We present biometric data for juve- niles (at the post-fledging stage) and adults. The mean wing length of juveniles, just before departure from the breeding grounds in mid June, reached 96% of that of adults. Juvenile total head lengths were 91% of adult, and bill and nalospi lengths 85% of adult, but tarsus and tarsus plus toe lengths were the same as adults. -

Behaviour of an Incubating Woodcock G
Behaviour of an incubating Woodcock G. des Forges INTRODUCTION In his well-known paper on the breeding habits of the'Woodcock Scolopax rusticola, Steinfatt (1938) records: 'The brooding female only rarely changes her position during the day; she lies for hours on the nest motionless. There seems to be a sort of rigidity, which overcomes the female. It obviously serves the purpose to reduce smell and so the possibility of being observed. Only twice a day, in morning and evening twilight, the female leaves the nest, in order to find food, for a total time of an hour'. A report on the European Woodcock (Shorten 1974) states that 'Steinfatt's description of behaviour at the nest seems to have been the basis for many subse quent accounts'. Also Vesey-Fitzgerald (1946), writing of his own experience in Surrey, says, 'I do not think that, unless disturbed, a sitting Woodcock leaves the nest during the day'. As circumstantial evidence had led me to believe that a sitting Woodcock did leave the nest and feed by day, I decided to attempt a prolonged watch on an incubating bird. THE NEST SITE The nest was in woodland, about 5 km north of Haywards Heath, West Sussex, on a hill-side sloping down from the main London to Brighton railway line to a stream at the bottom of the valley. The section of the wood concerned had been cleared of undergrowth and mature ash Fraxinus excelsior in 1972/73 leaving only standard oaks Quercus. Re-planting with mixed conifers had taken place in 1973/74 m tne open areas but not immediately round the nest, which was under the canopy of a group of six mature oaks, the lowest branches being 5 or 6 metres from the ground which here carried a thin growth of brambles Rubus fruticosus and bracken Pteridium aquilinum: but around the small conifers were only short grasses and a variety of perennials which had not made much growth by the end of March.