Simulating Current Regional Pattern and Composition of Chilean Native Forests Using a Dynamic Ecosystem Model
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Chapter14.Pdf
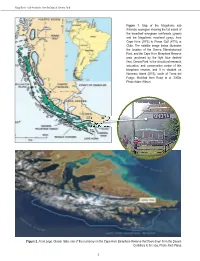
PART I • Omora Park Long-Term Ornithological Research Program THE OMORA PARK LONG-TERM ORNITHOLOGICAL RESEARCH PROGRAM: 1 STUDY SITES AND METHODS RICARDO ROZZI, JAIME E. JIMÉNEZ, FRANCISCA MASSARDO, JUAN CARLOS TORRES-MURA, AND RAJAN RIJAL In January 2000, we initiated a Long-term Ornithological Research Program at Omora Ethnobotanical Park in the world's southernmost forests: the sub-Antarctic forests of the Cape Horn Biosphere Reserve. In this chapter, we first present some key climatic, geographical, and ecological attributes of the Magellanic sub-Antarctic ecoregion compared to subpolar regions of the Northern Hemisphere. We then describe the study sites at Omora Park and other locations on Navarino Island and in the Cape Horn Biosphere Reserve. Finally, we describe the methods, including censuses, and present data for each of the bird species caught in mist nets during the first eleven years (January 2000 to December 2010) of the Omora Park Long-Term Ornithological Research Program. THE MAGELLANIC SUB-ANTARCTIC ECOREGION The contrast between the southwestern end of South America and the subpolar zone of the Northern Hemisphere allows us to more clearly distinguish and appreciate the peculiarities of an ecoregion that until recently remained invisible to the world of science and also for the political administration of Chile. So much so, that this austral region lacked a proper name, and it was generally subsumed under the generic name of Patagonia. For this reason, to distinguish it from Patagonia and from sub-Arctic regions, in the early 2000s we coined the name “Magellanic sub-Antarctic ecoregion” (Rozzi 2002). The Magellanic sub-Antarctic ecoregion extends along the southwestern margin of South America between the Gulf of Penas (47ºS) and Horn Island (56ºS) (Figure 1). -

Bioclimatic and Phytosociological Diagnosis of the Species of the Nothofagus Genus (Nothofagaceae) in South America
International Journal of Geobotanical Research, Vol. nº 1, December 2011, pp. 1-20 Bioclimatic and phytosociological diagnosis of the species of the Nothofagus genus (Nothofagaceae) in South America Javier AMIGO(1) & Manuel A. RODRÍGUEZ-GUITIÁN(2) (1) Laboratorio de Botánica, Facultad de Farmacia, Universidad de Santiago de Compostela (USC). E-15782 Santiago de Com- postela (Galicia, España). Phone: 34-881 814977. E-mail: [email protected] (2) Departamento de Producción Vexetal. Escola Politécnica Superior de Lugo-USC. 27002-Lugo (Galicia, España). E-mail: [email protected] Abstract The Nothofagus genus comprises 10 species recorded in the South American subcontinent. All are important tree species in the ex- tratropical, Mediterranean, temperate and boreal forests of Chile and Argentina. This paper presents a summary of data on the phyto- coenotical behaviour of these species and relates the plant communities to the measurable or inferable thermoclimatic and ombrocli- matic conditions which affect them. Our aim is to update the phytosociological knowledge of the South American temperate forests and to assess their suitability as climatic bioindicators by analysing the behaviour of those species belonging to their most represen- tative genus. Keywords: Argentina, boreal forests, Chile, mediterranean forests, temperate forests. Introduction tually give rise to a temperate territory with rainfall rates as high as those of regions with a Tropical pluvial bio- The South American subcontinent is usually associa- climate; iii. finally, towards the apex of the American ted with a tropical environment because this is in fact the Southern Cone, this temperate territory progressively dominant bioclimatic profile from Panamá to the north of gives way to a strip of land with a Boreal bioclimate. -

The Volcanic Ash Soils of Chile
' I EXPANDED PROGRAM OF TECHNICAL ASSISTANCE No. 2017 Report to the Government of CHILE THE VOLCANIC ASH SOILS OF CHILE FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS ROMEM965 -"'^ .Y--~ - -V^^-.. -r~ ' y Report No. 2017 Report CHT/TE/LA Scanned from original by ISRIC - World Soil Information, as ICSU World Data Centre for Soils. The purpose is to make a safe depository for endangered documents and to make the accrued information available for consultation, following Fair Use Guidelines. Every effort is taken to respect Copyright of the materials within the archives where the identification of the Copyright holder is clear and, where feasible, to contact the originators. For questions please contact [email protected] indicating the item reference number concerned. REPORT TO THE GOVERNMENT OP CHILE on THE VOLCANIC ASH SOILS OP CHILE Charles A. Wright POOL ANL AGRICULTURE ORGANIZATION OP THE UNITEL NATIONS ROME, 1965 266I7/C 51 iß - iii - TABLE OP CONTENTS Page INTRODUCTION 1 ACKNOWLEDGEMENTS 1 RECOMMENDATIONS 1 BACKGROUND INFORMATION 3 The nature and composition of volcanic landscapes 3 Vbloanio ash as a soil forming parent material 5 The distribution of voloanic ash soils in Chile 7 Nomenclature used in this report 11 A. ANDOSOLS OF CHILE» GENERAL CHARACTERISTICS, FORMATIVE ENVIRONMENT, AND MAIN KINDS OF SOIL 11 1. TRUMAO SOILS 11 General characteristics 11 The formative environment 13 ÈS (i) Climate 13 (ii) Topography 13 (iii) Parent materials 13 (iv) Natural plant cover 14 (o) The main kinds of trumao soils ' 14 2. NADI SOILS 16 General characteristics 16 The formative environment 16 tö (i) Climat* 16 (ii) Topograph? and parent materials 17 (iii) Natural plant cover 18 B. -

Pudu in a Chilean National Park
547 Pudu in a Chilean National Park Gary 8. Wetterberg The Chilean pudu Pudu pudu, the smallest American deer, is on the world list of endangered species in the IUCN Red Data Book. One of its few remaining refuges is in the Vicente Perez Rosales National Park. This is in the Lake District of southern Chile, the 'Switzerland of South America', between the Puyehue National Park to the north, and the Nahuel Huapi National Park in Argentina on the east. There are very few records on the fauna of this park, which covers 243,000 hectares, and is part of the Patagonian Subdivision of the Neotropical Faunal Region. Like an Island In many ways, Chile is like an island, cut off by the Atacama Desert on the north, the Andes to the east, the Patagonian ice fields and fiords to the south, and the Pacific on the west. This geo- graphical isolation has permitted the development of a unique biota, and Chilean wildlife exhibits some of the characteristics of island fauna such as narrow endemics and few competitors. The pudu is descended from the deer that migrated from North America in the late Tertiary period (Simpson 1950). The species is primarily of Chilean origin and distribution, although it is frequently encountered in adjacent areas of Argentina, and is present in Bolivia (Walker, 1964). It was discovered and named in 1782 by the Jesuit Juan Ignacio Molina, the 'father of Chilean natural history' (Osgood, 1943). Other species of the genus are found in Ecuador and Peru (Grimwood, 1968), and Brazil (Hershkovitz, 1958). -

Impact of Extreme Weather Events on Aboveground Net Primary Productivity and Sheep Production in the Magellan Region, Southernmost Chilean Patagonia
geosciences Article Impact of Extreme Weather Events on Aboveground Net Primary Productivity and Sheep Production in the Magellan Region, Southernmost Chilean Patagonia Pamela Soto-Rogel 1,* , Juan-Carlos Aravena 2, Wolfgang Jens-Henrik Meier 1, Pamela Gross 3, Claudio Pérez 4, Álvaro González-Reyes 5 and Jussi Griessinger 1 1 Institute of Geography, Friedrich–Alexander-University of Erlangen–Nürnberg, 91054 Erlangen, Germany; [email protected] (W.J.-H.M.); [email protected] (J.G.) 2 Centro de Investigación Gaia Antártica, Universidad de Magallanes, Punta Arenas 6200000, Chile; [email protected] 3 Servicio Agrícola y Ganadero (SAG), Punta Arenas 6200000, Chile; [email protected] 4 Private Consultant, Punta Arenas 6200000, Chile; [email protected] 5 Hémera Centro de Observación de la Tierra, Escuela de Ingeniería Forestal, Facultad de Ciencias, Universidad Mayor, Camino La Pirámide 5750, Huechuraba, Santiago 8580745, Chile; [email protected] * Correspondence: [email protected] Received: 28 June 2020; Accepted: 13 August 2020; Published: 16 August 2020 Abstract: Spatio-temporal patterns of climatic variability have effects on the environmental conditions of a given land territory and consequently determine the evolution of its productive activities. One of the most direct ways to evaluate this relationship is to measure the condition of the vegetation cover and land-use information. In southernmost South America there is a limited number of long-term studies on these matters, an incomplete network of weather stations and almost no database on ecosystems productivity. In the present work, we characterized the climate variability of the Magellan Region, southernmost Chilean Patagonia, for the last 34 years, studying key variables associated with one of its main economic sectors, sheep production, and evaluating the effect of extreme weather events on ecosystem productivity and sheep production. -

Chile: a Journey to the End of the World in Search of Temperate Rainforest Giants
Eliot Barden Kew Diploma Course 53 July 2017 Chile: A Journey to the end of the world in search of Temperate Rainforest Giants Valdivian Rainforest at Alerce Andino Author May 2017 1 Eliot Barden Kew Diploma Course 53 July 2017 Table of Contents 1. Title Page 2. Contents 3. Table of Figures/Introduction 4. Introduction Continued 5. Introduction Continued 6. Aims 7. Aims Continued / Itinerary 8. Itinerary Continued / Objective / the Santiago Metropolitan Park 9. The Santiago Metropolitan Park Continued 10. The Santiago Metropolitan Park Continued 11. Jardín Botánico Chagual / Jardin Botanico Nacional, Viña del Mar 12. Jardin Botanico Nacional Viña del Mar Continued 13. Jardin Botanico Nacional Viña del Mar Continued 14. Jardin Botanico Nacional Viña del Mar Continued / La Campana National Park 15. La Campana National Park Continued / Huilo Huilo Biological Reserve Valdivian Temperate Rainforest 16. Huilo Huilo Biological Reserve Valdivian Temperate Rainforest Continued 17. Huilo Huilo Biological Reserve Valdivian Temperate Rainforest Continued 18. Huilo Huilo Biological Reserve Valdivian Temperate Rainforest Continued / Volcano Osorno 19. Volcano Osorno Continued / Vicente Perez Rosales National Park 20. Vicente Perez Rosales National Park Continued / Alerce Andino National Park 21. Alerce Andino National Park Continued 22. Francisco Coloane Marine Park 23. Francisco Coloane Marine Park Continued 24. Francisco Coloane Marine Park Continued / Outcomes 25. Expenditure / Thank you 2 Eliot Barden Kew Diploma Course 53 July 2017 Table of Figures Figure 1.) Valdivian Temperate Rainforest Alerce Andino [Photograph; Author] May (2017) Figure 2. Map of National parks of Chile Figure 3. Map of Chile Figure 4. Santiago Metropolitan Park [Photograph; Author] May (2017) Figure 5. -

New Jan16.2011
Spring 2011 Mail Order Catalog Cistus Nursery 22711 NW Gillihan Road Sauvie Island, OR 97231 503.621.2233 phone 503.621.9657 fax order by phone 9 - 5 pst, visit 10am - 5pm, fax, mail, or email: [email protected] 24-7-365 www.cistus.com Spring 2011 Mail Order Catalog 2 USDA zone: 2 Symphoricarpos orbiculatus ‘Aureovariegatus’ coralberry Old fashioned deciduous coralberry with knock your socks off variegation - green leaves with creamy white edges. Pale white-tinted-pink, mid-summer flowers attract bees and butterflies and are followed by bird friendly, translucent, coral berries. To 6 ft or so in most any normal garden conditions - full sun to part shade with regular summer water. Frost hardy in USDA zone 2. $12 Caprifoliaceae USDA zone: 3 Athyrium filix-femina 'Frizelliae' Tatting fern An unique and striking fern with narrow fronds, only 1" wide and oddly bumpy along the sides as if beaded or ... tatted. Found originally in the Irish garden of Mrs. Frizell and loved for it quirkiness ever since. To only 1 ft tall x 2 ft wide and deciduous, coming back slowly in spring. Best in bright shade or shade where soil is rich. Requires summer water. Frost hardy to -40F, USDA zone 3 and said to be deer resistant. $14 Woodsiaceae USDA zone: 4 Aralia cordata 'Sun King' perennial spikenard The foliage is golden, often with red stems, and dazzling on this big and bold perennial, quickly to 3 ft tall and wide, first discovered in a department store in Japan by nurseryman Barry Yinger. Spikes of aralia type white flowers in summer are followed by purple-black berries. -

Bosque Pehuén Park's Flora: a Contribution to the Knowledge of the Andean Montane Forests in the Araucanía Region, Chile Author(S): Daniela Mellado-Mansilla, Iván A
Bosque Pehuén Park's Flora: A Contribution to the Knowledge of the Andean Montane Forests in the Araucanía Region, Chile Author(s): Daniela Mellado-Mansilla, Iván A. Díaz, Javier Godoy-Güinao, Gabriel Ortega-Solís and Ricardo Moreno-Gonzalez Source: Natural Areas Journal, 38(4):298-311. Published By: Natural Areas Association https://doi.org/10.3375/043.038.0410 URL: http://www.bioone.org/doi/full/10.3375/043.038.0410 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. R E S E A R C H A R T I C L E ABSTRACT: In Chile, most protected areas are located in the southern Andes, in mountainous land- scapes at mid or high altitudes. Despite the increasing proportion of protected areas, few have detailed inventories of their biodiversity. This information is essential to define threats and develop long-term • integrated conservation programs to face the effects of global change. -

Bioclimatic and Phytosociological Diagnosis of the Species of the Nothofagus Genus (Nothofagaceae) in South America
International Journal of Geobotanical Research, Vol. nº 1, December 2011, pp. 1-20 Bioclimatic and phytosociological diagnosis of the species of the Nothofagus genus (Nothofagaceae) in South America Javier AMIGO(1) & Manuel A. RODRÍGUEZ-GUITIÁN(2) (1) Laboratorio de Botánica, Facultad de Farmacia, Universidad de Santiago de Compostela (USC). E-15782 Santiago de Com- postela (Galicia, España). Phone: 34-881 814977. E-mail: [email protected] (2) Departamento de Producción Vexetal. Escola Politécnica Superior de Lugo-USC. 27002-Lugo (Galicia, España). E-mail: [email protected] Abstract The Nothofagus genus comprises 10 species recorded in the South American subcontinent. All are important tree species in the ex- tratropical, Mediterranean, temperate and boreal forests of Chile and Argentina. This paper presents a summary of data on the phyto- coenotical behaviour of these species and relates the plant communities to the measurable or inferable thermoclimatic and ombrocli- matic conditions which affect them. Our aim is to update the phytosociological knowledge of the South American temperate forests and to assess their suitability as climatic bioindicators by analysing the behaviour of those species belonging to their most represen- tative genus. Keywords: Argentina, boreal forests, Chile, mediterranean forests, temperate forests. Introduction tually give rise to a temperate territory with rainfall rates as high as those of regions with a Tropical pluvial bio- The South American subcontinent is usually associa- climate; iii. finally, towards the apex of the American ted with a tropical environment because this is in fact the Southern Cone, this temperate territory progressively dominant bioclimatic profile from Panamá to the north of gives way to a strip of land with a Boreal bioclimate. -

Shoot Development and Dieback in Progenies of Nothofagus Obliqua Javier G
Shoot development and dieback in progenies of Nothofagus obliqua Javier G. Puntieri, Javier E. Grosfeld, Marina Stecconi, Cecilia Brion, María Marta Azpilicueta, Leonardo Gallo, Daniel Barthélémy To cite this version: Javier G. Puntieri, Javier E. Grosfeld, Marina Stecconi, Cecilia Brion, María Marta Azpilicueta, et al.. Shoot development and dieback in progenies of Nothofagus obliqua. Annals of Forest Science, Springer Nature (since 2011)/EDP Science (until 2010), 2007, 64 (8), pp.839-844. 10.1051/forest:2007068. hal-00259241 HAL Id: hal-00259241 https://hal.archives-ouvertes.fr/hal-00259241 Submitted on 30 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Copyright Ann. For. Sci. 64 (2007) 839–844 Available online at: c INRA, EDP Sciences, 2007 www.afs-journal.org DOI: 10.1051/forest:2007068 Original article Shoot development and dieback in progenies of Nothofagus obliqua Javier Puntieria,b*,JavierGrosfelda,b,MarinaStecconib, Cecilia Briona, María Marta Azpilicuetac, Leonardo Galloc,DanielBarthel´ emy´ d a Departamento de Botánica, Universidad Nacional del Comahue, Quintral 1250, 8400, Bariloche, Argentina b Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina c Laboratorio de Genética Forestal, Instituto Nacional de Tecnología Agropecuaria, EEA Bariloche, Argentina d INRA, Unité Mixte de Recherche CIRAD-CNRS-INRA-IRD-Université Montpellier 2, “ botAnique et bioinforMatique de l’Architecture des Plantes ” (AMAP), UMR T51 (CIRAD), UMR 5120 (CNRS), UMR 931 (INRA), M123 (IRD), UM27 (UMII) TA A-51/PS2, Blvd. -

Actualización De La Clasificación De Tipos Forestales Y Cobertura Del Suelo De La Región Bosque Andino Patagónico
ACTUALIZACIÓN DE LA CLASIFICACIÓN DE TIPOS FORESTALES Y COBERTURA DEL SUELO DE LA REGIÓN BOSQUE ANDINO PATAGÓNICO INFORME FINAL Julio 2016 CARTOGRAFÍA PARA EL INVENTARIO FORESTAL NACIONAL DE BOSQUES NATIVOS Mapa base para un sistema de monitoreo continuo de la región Información de base 2013 - 2014 Cita recomendada de esta versión del trabajo: CIEFAP, MAyDS, 2016. Actualización de la Clasificación de Tipos Forestales y Cobertura del Suelo de la Región Bosque Andino Patagónico. Informe Final. CIEFAP. https://drive.google.com/open?id=0BxfNQUtfxxeaUHNCQm9lYmk5RnM 2 Presidente de la Nación Ing. Mauricio Macri Ministro de Ambiente y Desarrollo Sustentable de la Nación Rabino Sergio Alejandro Bergman Secretario de Política Ambiental, Cambio Climático y Desarrollo Sustentable Lic. Diego Moreno Sub Secretaria de Planificación y Ordenamiento del Territorio Dra. Dolores Duverges Directora Nacional de Bosques, Ordenamiento Territorial y Suelos Dra. María Esperanza Alonso Director de Bosques Ing. Rubén Manfredi 3 Dirección General de Recursos Forestales de la Provincia de Neuquén Téc. Ftal. Uriel Mele Subsecretaría de Recursos Forestales de la Provincia de Río Negro Ing. Marcelo Perdomo Subsecretaría de Bosques de la Provincia del Chubut Sr. Leonardo Aquilanti Dirección de Bosques de la Provincia de Santa Cruz Ing. Julia Chazarreta Dirección General de Bosques Secretaría de Ambiente, Desarrollo Sustentable y Cambio Climático Provincia de Tierra del Fuego, Antártida e Islas del Atlántico Sur Ing. Gustavo Cortés Administración Parques Nacionales Sr. Eugenio Breard 4 Este trabajo fue realizado por el Nodo Regional Bosque Andino Patagónico (BAP) con la participación de las jurisdicciones regionales. El Nodo BAP tiene sede en el Centro de Investigación y Extensión Forestal Andino Patagónico (CIEFAP) Durante la ejecución de este proyecto, lamentablemente nos dejó de manera trágica nuestro amigo y compañero de trabajo, Ing. -

The Pine Genome Initiative- Science Plan Review
ProCoGen Training Workshop 2013 Umeå An Undiscovered Country: What Comparative Genomics Tells Us of Gymnosperm Genomes Ujwal R. Bagal, W. Walter Lorenz, Jeffrey F.D. Dean Warnell School of Forestry and Natural Resources & Institute of Bioinformatics University of Georgia Diverse Form and Life History JGI CSP - Conifer EST Project Transcriptome Assemblies Statistics Pinaceae Reads Contigs* • Pinus taeda 4,074,360 164,506 • Pinus palustris 542,503 44,975 • Pinus lambertiana 1,096,017 85,348 • Picea abies 623,144 36,867 • Cedrus atlantica 408,743 30,197 • Pseudotsuga menziesii 1,216,156 60,504 Other Conifer Families • Wollemia nobilis 471,719 35,814 • Cephalotaxus harringtonia 689,984 40,884 • Sequoia sempervirens 472,601 42,892 • Podocarpus macrophylla 584,579 36,624 • Sciadopitys verticillata 479,239 29,149 • Taxus baccata 398,037 33,142 *Assembled using MIRA http://ancangio.uga.edu/ng-genediscovery/conifer_dbMagic.jnlp Loblolly pine PAL amino acid sequence alignment Analysis Method Sequence Collection PlantTribe, PlantGDB, GenBank, Conifer DBMagic assemblies 25 taxa comprising of 71 sequences Phylogenetic analysis Maximum Likelihood: RAxML (Stamatakis et. al) Bayesian Method: MrBayes (Huelsenbeck, et al.) Tree reconciliation: NOTUNG 2.6 (Chen et al.) Phylogenetic Tree of Vascular Plant PALs Phylogenetic Analysis of Conifer PAL Gene Sequences Conifer-specific branch shown in green Amino Acids Under Relaxed Constraint Maximum Likelihood analysis Nested codon substitution models M0 : constant dN/dS ratio M2a : rate ratio ω1< 1, ω2=1 and ω3>1 M3 : (ω1< ω2< ω3) (M0, M2a, M3, M2a+S1, M2a+S2, M3+S1, M3+S2) Fitmodel version 0.5.3 ( Guindon et al. 2004) S1 : equal switching rates (alpha =beta) S2 : unequal switching rates (alpha ≠ beta) Variable gymnosperm PAL amino acid residues mapped onto a crystal structure for parsley PAL Ancestral polyploidy in seed plants and angiosperms Jiao et al.