Bosque Pehuén Park's Flora: a Contribution to the Knowledge of the Andean Montane Forests in the Araucanía Region, Chile Author(S): Daniela Mellado-Mansilla, Iván A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Toward a Resolution of Campanulid Phylogeny, with Special Reference to the Placement of Dipsacales
TAXON 57 (1) • February 2008: 53–65 Winkworth & al. • Campanulid phylogeny MOLECULAR PHYLOGENETICS Toward a resolution of Campanulid phylogeny, with special reference to the placement of Dipsacales Richard C. Winkworth1,2, Johannes Lundberg3 & Michael J. Donoghue4 1 Departamento de Botânica, Instituto de Biociências, Universidade de São Paulo, Caixa Postal 11461–CEP 05422-970, São Paulo, SP, Brazil. [email protected] (author for correspondence) 2 Current address: School of Biology, Chemistry, and Environmental Sciences, University of the South Pacific, Private Bag, Laucala Campus, Suva, Fiji 3 Department of Phanerogamic Botany, The Swedish Museum of Natural History, Box 50007, 104 05 Stockholm, Sweden 4 Department of Ecology & Evolutionary Biology and Peabody Museum of Natural History, Yale University, P.O. Box 208106, New Haven, Connecticut 06520-8106, U.S.A. Broad-scale phylogenetic analyses of the angiosperms and of the Asteridae have failed to confidently resolve relationships among the major lineages of the campanulid Asteridae (i.e., the euasterid II of APG II, 2003). To address this problem we assembled presently available sequences for a core set of 50 taxa, representing the diver- sity of the four largest lineages (Apiales, Aquifoliales, Asterales, Dipsacales) as well as the smaller “unplaced” groups (e.g., Bruniaceae, Paracryphiaceae, Columelliaceae). We constructed four data matrices for phylogenetic analysis: a chloroplast coding matrix (atpB, matK, ndhF, rbcL), a chloroplast non-coding matrix (rps16 intron, trnT-F region, trnV-atpE IGS), a combined chloroplast dataset (all seven chloroplast regions), and a combined genome matrix (seven chloroplast regions plus 18S and 26S rDNA). Bayesian analyses of these datasets using mixed substitution models produced often well-resolved and supported trees. -
Chapter14.Pdf
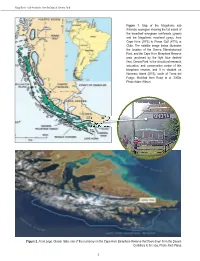
PART I • Omora Park Long-Term Ornithological Research Program THE OMORA PARK LONG-TERM ORNITHOLOGICAL RESEARCH PROGRAM: 1 STUDY SITES AND METHODS RICARDO ROZZI, JAIME E. JIMÉNEZ, FRANCISCA MASSARDO, JUAN CARLOS TORRES-MURA, AND RAJAN RIJAL In January 2000, we initiated a Long-term Ornithological Research Program at Omora Ethnobotanical Park in the world's southernmost forests: the sub-Antarctic forests of the Cape Horn Biosphere Reserve. In this chapter, we first present some key climatic, geographical, and ecological attributes of the Magellanic sub-Antarctic ecoregion compared to subpolar regions of the Northern Hemisphere. We then describe the study sites at Omora Park and other locations on Navarino Island and in the Cape Horn Biosphere Reserve. Finally, we describe the methods, including censuses, and present data for each of the bird species caught in mist nets during the first eleven years (January 2000 to December 2010) of the Omora Park Long-Term Ornithological Research Program. THE MAGELLANIC SUB-ANTARCTIC ECOREGION The contrast between the southwestern end of South America and the subpolar zone of the Northern Hemisphere allows us to more clearly distinguish and appreciate the peculiarities of an ecoregion that until recently remained invisible to the world of science and also for the political administration of Chile. So much so, that this austral region lacked a proper name, and it was generally subsumed under the generic name of Patagonia. For this reason, to distinguish it from Patagonia and from sub-Arctic regions, in the early 2000s we coined the name “Magellanic sub-Antarctic ecoregion” (Rozzi 2002). The Magellanic sub-Antarctic ecoregion extends along the southwestern margin of South America between the Gulf of Penas (47ºS) and Horn Island (56ºS) (Figure 1). -

From Witch Doctor to Modern Medicine: Searching the American Tropics for Potentially New Medicinal Plants
From Witch Doctor to Modern Medicine: Searching the American Tropics for Potentially New Medicinal Plants "And as there are discouvered new Regions, new King- doms and new Provinces by our Spaniards, so they have brought unto us new Medicines, and newe Remedies, wherewith they do cure many infirmities, which, if we did lacke them, would be incurible, and without any remedie ... for which cause I did pretend to treate, and to write of all things that they bring from our Indias, apperteyning to the Arte and Use of Medicine, and the remedie of hurtes and diseases that we doo suffer and endure ...". Nicholas Monardes [transl. John Frampton] "Joyfull Newes out of the New-found World" (1596) The search for potential new medicines from the Plant King- dom is not new. It has been carried on since man first became aware ot the variety ot ettects that plants had on his mind and body. It is still under way. But we now have many ad- vantages over man in all prior ages. We have millennia of experience behind us; a deeper understanding of what plants are and how they live; an intensity of modern technology to help us; and, what is more important than all of the other advantages, a determination to probe into the still virgin vegetal world for active compounds of promise for man’s existence. Today we follow several discreet paths in our search for "new" medicinal plants. Some modern programs - ambitious, technologically com- plex and potentially promising - have emphasized the random or semi-random screening of plants from all parts of the world for new organic compounds. -

NSW Rainforest Trees Part
This document has been scanned from hard-copy archives for research and study purposes. Please note not all information may be current. We have tried, in preparing this copy, to make the content accessible to the widest possible audience but in some cases we recognise that the automatic text recognition maybe inadequate and we apologise in advance for any inconvenience this may cause. · RESEARCH NOTE No. 35 ~.I~=1 FORESTRY COMMISSION OF N.S.W. RESEARCH NOTE No. 35 P)JBLISHED 197R N.S.W. RAINFOREST TREES PART VII FAMILIES: PROTEACEAE SANTALACEAE NYCTAGINACEAE GYROSTEMONACEAE ANNONACEAE EUPOMATIACEAE MONIMIACEAE AUTHOR A.G.FLOYD (Research Note No. 35) National Library of Australia card number and ISBN ISBN 0 7240 13997 ISSN 0085-3984 INTRODUCTION This is the seventh in a series ofresearch notes describing the rainforest trees of N.S. W. Previous publications are:- Research Note No. 3 (I 960)-N.S.W. Rainforest Trees. Part I Family LAURACEAE. A. G. Floyd and H. C. Hayes. Research Note No. 7 (1961)-N.S.W. Rainforest Trees. Part II Families Capparidaceae, Escalloniaceae, Pittosporaceae, Cunoniaceae, Davidsoniaceae. A. G. Floyd and H. C. Hayes. Research Note No. 28 (I 973)-N.S.W. Rainforest Trees. Part III Family Myrtaceae. A. G. Floyd. Research Note No. 29 (I 976)-N.S.W. Rainforest Trees. Part IV Family Rutaceae. A. G. Floyd. Research Note No. 32 (I977)-N.S.W. Rainforest Trees. Part V Families Sapindaceae, Akaniaceae. A. G. Floyd. Research Note No. 34 (1977)-N.S.W. Rainforest Trees. Part VI Families Podocarpaceae, Araucariaceae, Cupressaceae, Fagaceae, Ulmaceae, Moraceae, Urticaceae. -

PROTEACEAE – It's All About Pollination
PROTEACEAE – it’s all about pollination …….Gail Slykhuis Illustration Philippa Hesterman, images Ellinor Campbell & Marg McDonald A predominantly southern hemisphere plant family, Proteaceae is well represented in Australia, particularly in the West, but we do have our own equally special local representatives, some of which are outlined below. A characteristic feature of many genera within this plant family is the ‘pollen presenter’, which is a fascinating mechanism by which the pollen, which would otherwise be difficult to access for potential pollination vectors such as bees, birds and nectarivorous mammals, is positioned on the extended style of the flower, facilitating cross- pollination. The stigma, which is part of the style, is not mature at this time, thus avoiding self-pollination. A hand lens would enable you to clearly see pollen presenters on the following local representatives: Banksia marginata, Grevillea infecunda, Hakea spp., Isopogon ceratophyllus and Lomatia illicifolia. It is interesting to note that both Victorian Smoke-bush Conospermum mitchellii and Prickly Geebung Persoonia juniperina, also found in our district, do not have pollen presenters. Silver Banksia Banksia marginata This shrub or small tree is readily recognisable when flowering (Feb – July) by the conspicuous yellow pollen presenters, which are an obvious floral part of the banksia flower. These flowers then slowly mature into our iconic woody banksia cones. It is interesting to observe the changes in the nature of the pollen presenters as the flower develops. The white undersides of the leathery leaves provide a clue to the choice of common name with their tip being characteristically blunt or truncate. Anglesea Grevillea Grevillea infecunda One of our endemic plants, the Anglesea Grevillea was first named in 1986 and is Anglesea Grevillea found in several locations north west of Anglesea. -

N E W S L E T T E R
N E W S L E T T E R PLANTS OF TASMANIA Nursery and Gardens 65 Hall St Ridgeway TAS 7054 Open 7 Days a week – 9 am to 5 pm Closed Christmas Day, Boxing Day and Good Friday Phone: (03) 6239 1583 Fax: (03) 6239 1106 Email: [email protected] Newsletter 26 Spring 2011 Website: www.potn.com.au Hello, and welcome to the spring newsletter for 2011! News from the Nursery We are madly propagating at the moment, with many thousands of new cuttings putting their roots out and seedlings popping their heads up above the propagating mix. It is always an exciting time, as we experiment with seed from new species – sometimes they work, and sometimes we understand why we’ve never grown them before... New plants should start being put out into the sales area soon – fresh-faced little things ready to pop into the ground! We have recently purchased a further block of land from the ex-neighbours Jubilee Nursery, now sadly closed, that will give us a lot more flexibility and the ability to grow and store more plants. As mentioned last newsletter we have done some major revamping in the garden. A lot of work by all the staff has led to a much more open garden with a lovely Westringia brevifolia hedge (well, it will be a hedge when it grows a bit), another Micrantheum hexandrum Cream Cascade hedge-to-be, lots of Correas, Lomatias and Baueras. Where we sell a few forms of a particular species we have tried to plant examples of each so that we can show you what they are like. -

Appendix 1: Maps and Plans Appendix184 Map 1: Conservation Categories for the Nominated Property
Appendix 1: Maps and Plans Appendix184 Map 1: Conservation Categories for the Nominated Property. Los Alerces National Park, Argentina 185 Map 2: Andean-North Patagonian Biosphere Reserve: Context for the Nominated Proprty. Los Alerces National Park, Argentina 186 Map 3: Vegetation of the Valdivian Ecoregion 187 Map 4: Vegetation Communities in Los Alerces National Park 188 Map 5: Strict Nature and Wildlife Reserve 189 Map 6: Usage Zoning, Los Alerces National Park 190 Map 7: Human Settlements and Infrastructure 191 Appendix 2: Species Lists Ap9n192 Appendix 2.1 List of Plant Species Recorded at PNLA 193 Appendix 2.2: List of Animal Species: Mammals 212 Appendix 2.3: List of Animal Species: Birds 214 Appendix 2.4: List of Animal Species: Reptiles 219 Appendix 2.5: List of Animal Species: Amphibians 220 Appendix 2.6: List of Animal Species: Fish 221 Appendix 2.7: List of Animal Species and Threat Status 222 Appendix 3: Law No. 19,292 Append228 Appendix 4: PNLA Management Plan Approval and Contents Appendi242 Appendix 5: Participative Process for Writing the Nomination Form Appendi252 Synthesis 252 Management Plan UpdateWorkshop 253 Annex A: Interview Guide 256 Annex B: Meetings and Interviews Held 257 Annex C: Self-Administered Survey 261 Annex D: ExternalWorkshop Participants 262 Annex E: Promotional Leaflet 264 Annex F: Interview Results Summary 267 Annex G: Survey Results Summary 272 Annex H: Esquel Declaration of Interest 274 Annex I: Trevelin Declaration of Interest 276 Annex J: Chubut Tourism Secretariat Declaration of Interest 278 -

Lições Das Interações Planta – Beija-Flor
UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE BIOLOGIA JÉFERSON BUGONI REDES PLANTA-POLINIZADOR NOS TRÓPICOS: LIÇÕES DAS INTERAÇÕES PLANTA – BEIJA-FLOR PLANT-POLLINATOR NETWORKS IN THE TROPICS: LESSONS FROM HUMMINGBIRD-PLANT INTERACTIONS CAMPINAS 2017 JÉFERSON BUGONI REDES PLANTA-POLINIZADOR NOS TRÓPICOS: LIÇÕES DAS INTERAÇÕES PLANTA – BEIJA-FLOR PLANT-POLLINATOR NETWORKS IN THE TROPICS: LESSONS FROM HUMMINGBIRD-PLANT INTERACTIONS Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do Título de Doutor em Ecologia. Thesis presented to the Institute of Biology of the University of Campinas in partial fulfillment of the requirements for the degree of Doctor in Ecology. ESTE ARQUIVO DIGITAL CORRESPONDE À VERSÃO FINAL DA TESE DEFENDIDA PELO ALUNO JÉFERSON BUGONI E ORIENTADA PELA DRA. MARLIES SAZIMA. Orientadora: MARLIES SAZIMA Co-Orientador: BO DALSGAARD CAMPINAS 2017 Campinas, 17 de fevereiro de 2017. COMISSÃO EXAMINADORA Profa. Dra. Marlies Sazima Prof. Dr. Felipe Wanderley Amorim Prof. Dr. Thomas Michael Lewinsohn Profa. Dra. Marina Wolowski Torres Prof. Dr. Vinícius Lourenço Garcia de Brito Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no processo de vida acadêmica do aluno. DEDICATÓRIA À minha família por me ensinar o amor à natureza e a natureza do amor. Ao povo brasileiro por financiar meus estudos desde sempre, fomentando assim meus sonhos. EPÍGRAFE “Understanding patterns in terms of the processes that produce them is the essence of science […]” Levin, S.A. (1992). The problem of pattern and scale in ecology. Ecology 73:1943–1967. AGRADECIMENTOS Manifestar a gratidão às tantas pessoas que fizeram parte direta ou indiretamente do processo que culmina nesta tese não é tarefa trivial. -

Principles and Practice of Forest Landscape Restoration Case Studies from the Drylands of Latin America Edited by A.C
Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor About IUCN IUCN, International Union for Conservation of Nature, helps the world find pragmatic solutions to our most pressing environment and development challenges. IUCN works on biodiversity, climate change, energy, human livelihoods and greening the world economy by supporting scientific research, managing field projects all over the world, and bringing governments, NGOs, the UN and companies together to develop policy, laws and best practice. IUCN is the world’s oldest and largest global environmental organization, with more than 1,000 government and NGO members and almost 11,000 volunteer experts in some 160 countries. IUCN’s work is supported by over 1,000 staff in 60 offices and hundreds of partners in public, NGO and private sectors around the world. www.iucn.org Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor This book is dedicated to the memory of Margarito Sánchez Carrada, a student who worked on the research project described in these pages. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN or the European Commission concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Systematics of Chusquea Section Chusquea, Section Swallenochloa, Section Verticillatae, and Section Serpentes (Poaceae: Bambusoideae) Lynn G
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1986 Systematics of Chusquea section Chusquea, section Swallenochloa, section Verticillatae, and section Serpentes (Poaceae: Bambusoideae) Lynn G. Clark Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Botany Commons Recommended Citation Clark, Lynn G., "Systematics of Chusquea section Chusquea, section Swallenochloa, section Verticillatae, and section Serpentes (Poaceae: Bambusoideae) " (1986). Retrospective Theses and Dissertations. 7988. https://lib.dr.iastate.edu/rtd/7988 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This reproduction was made from a copy of a manuscript sent to us for publication and microfilming. While the most advanced technology has been used to pho tograph and reproduce this manuscript, the quality of the reproduction is heavily dependent upon the quality of the material submitted. Pages in any manuscript may have indistinct print. In all cases the best available copy has been filmed. The following explanation of techniques Is provided to help clarify notations which may appear on this reproduction. 1. Manuscripts may not always be complete. When it is not possible to obtain missing jiages, a note appears to indicate this. 2. When copyrighted materials are removed from the manuscript, a note ap pears to indicate this. 3. -

Principles and Practice of Forest Landscape Restoration Case Studies from the Drylands of Latin America Edited by A.C
Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor About IUCN IUCN, International Union for Conservation of Nature, helps the world find pragmatic solutions to our most pressing environment and development challenges. IUCN works on biodiversity, climate change, energy, human livelihoods and greening the world economy by supporting scientific research, managing field projects all over the world, and bringing governments, NGOs, the UN and companies together to develop policy, laws and best practice. IUCN is the world’s oldest and largest global environmental organization, with more than 1,000 government and NGO members and almost 11,000 volunteer experts in some 160 countries. IUCN’s work is supported by over 1,000 staff in 60 offices and hundreds of partners in public, NGO and private sectors around the world. www.iucn.org Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor This book is dedicated to the memory of Margarito Sánchez Carrada, a student who worked on the research project described in these pages. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN or the European Commission concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Università Degli Studi Di Milano
UNIVERSITÀ DEGLI STUDI DI MILANO Dottorato in Scienze Farmacologiche Sperimentali e Cliniche XXXII ciclo Dipartimento di Scienze Farmacologiche e Biomolecolari VALIDATION OF PLANTS TRADITIONALLY USED FOR SKIN INFLAMMATION Settore Scientifico Disciplinare BIO/14 Saba KHALILPOUR Tutor: Prof. Mario DELL’AGLI Coordinatore: Prof. Alberico L. CATAPANO A.A. 2018 - 2019 1 TABLE OF CONTENTS TABLE OF CONTENTS .........................................................................................2 LIST OF ABBREVIATIONS ..................................................................................6 LIST OF SYMBOLS ...............................................................................................8 RIASSUNTO ...........................................................................................................9 ABSTRACT ...........................................................................................................12 CHAPTER ONE ....................................................................................................15 1. Introduction ...........................................................................................15 1.1 Basic structure and functions of the skin .........................................................15 1.1.1 Epidermis .................................................................................16 1.1.1.1 Keratinocytes ......................................................................18 1.1.1.2 Other cell types ..................................................................19