Breeding and Feeding Ecology of Bald Eagl~S in the Apostle Island National Lakeshore
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Great Lakes Islands: Biodiversity Elements And
GREAT LAKES ISLANDS: BIODIVERSITY ELEMENTS AND THREATS A FINAL REPORT TO THE GREAT LAKES NATIONAL PROGRAM OFFICE OF THE ENVIRONMENTAL PROTECTION AGENCY AUGUST 6, 2007 ACKNOWLEDGMENTS Funding for this project has been provided by the Great Lakes Program Office (GLNPO) of the Environmental Protection Agency (Grant No. Gl-96521901: Framework for the Binational Conservation of Great Lakes Islands). We especially appreciated the support of our project officer, K. Rodriquez, and G. Gulezian, director of the GLNPO. Project team members were F. Cuthbert (University of Minnesota), D. Ewert (The Nature Conservancy), R. Greenwood (U. S. Fish & Wildlife Service), D. Kraus (The Nature Conservancy of Canada), M. Seymour (U.S. Fish & Wildlife Service), K. Vigmostad (Principal Investigator, formerly of Northeast-Midwest Institute), and L. Wires (University of Minnesota). Team members for the Ontario portion of the project included W. Bakowsky (NHIC), B. Crins (Ontario Parks), J. Mackenzie (NHIC) and M. McMurtry (NHIC). GIS and technical support for this project has been provided by T. Krahn (Provincial Geomatics Service Centre, OMNR), J. Slatts (The Nature Conservancy), and G. White (The Nature Conservancy of Canada). Many others have provided scientific and policy support for this project. We particularly want to recognize M. DePhillips (The Nature Conservancy), G. Jackson (Parks Canada), B. Manny (Great Lakes Science Center), and C. Vasarhelyi (policy consultant). Cover photograph: A Bay on Gibraltar Island (Lake Erie) ©2005 Karen E. Vigmostad 2 Contents -
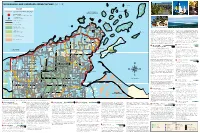
WLSSB Map and Guide
WISCONSIN LAKE SUPERIOR SCENIC BYWAY (WLSSB) DEVILS ISLAND NORTH TWIN ISLAND MAP KEY ROCKY ISLAND SOUTH TWIN ISLAND CAT ISLAND WISCONSIN LAKE SUPERIOR SCENIC BYWAY APOSTLE ISLANDS BEAR ISLAND NATIONAL LAKESHORE KIOSK LOCATION IRONWOOD ISLAND SCENIC BYWAY NEAR HERBSTER SAILING ON LAKE SUPERIOR LOST CREEK FALLS KIOSKS CONTAIN DETAILED INFORMATION ABOUT EACH LOCATION SAND ISLAND VISITOR INFORMATION OUTER ISLAND YORK ISLAND SEE REVERSE FOR COMPLETE LIST µ OTTER ISLAND FEDERAL HIGHWAY MANITOU ISLAND RASPBERRY ISLAND STATE HIGHWAY COUNTY HIGHWAY 7 EAGLE ISLAND NATIONAL PARKS ICE CAVES AT MEYERS BEACH BAYFIELD PENINSULA AND THE APOSTLE ISLANDS FROM MT. ASHWABAY & NATIONAL FOREST LANDS well as a Heritage Museum and a Maritime Museum. Pick up Just across the street is the downtown area with a kayak STATE PARKS K OAK ISLAND STOCKTON ISLAND some fresh or smoked fish from a commercial fishery for a outfitter, restaurants, more lodging and a historic general & STATE FOREST LANDS 6 GULL ISLAND taste of Lake Superior or enjoy local flavors at one of the area store that has a little bit of everything - just like in the “old (!13! RED CLIFF restaurants. If you’re brave, try the whitefish livers – they’re a days,” but with a modern flair. Just off the Byway you can MEYERS BEACH COUNTY PARKS INDIAN RESERVATION local specialty! visit two popular waterfalls: Siskiwit Falls and Lost Creek & COUNTY FOREST LANDS Falls. West of Cornucopia you will find the Lost Creek Bog HERMIT ISLAND Walk the Brownstone Trail along an old railroad grade or CORNUCOPIA State Natural Area. Lost Creek Bog forms an estuary at the take the Gil Larson Nature Trail (part of the Big Ravine Trail MICHIGAN ISLAND mouths of three small creeks (Lost Creek 1, 2, and 3) where System) which starts by a historic apple shed, continues RESERVATION LANDS they empty into Lake Superior at Siskiwit Bay. -

Apostle Islands National Lakehore Geologic Resources Inventory
Geologic Resources Inventory Scoping Summary Apostle Islands National Lakeshore Geologic Resources Division Prepared by Trista L. Thornberry-Ehrlich National Park Service August 7, 2010 US Department of the Interior The Geologic Resources Inventory (GRI) provides each of 270 identified natural area National Park System units with a geologic scoping meeting and summary (this document), a digital geologic map, and a geologic resources inventory report. The purpose of scoping is to identify geologic mapping coverage and needs, distinctive geologic processes and features, resource management issues, and monitoring and research needs. Geologic scoping meetings generate an evaluation of the adequacy of existing geologic maps for resource management, provide an opportunity to discuss park-specific geologic management issues, and if possible include a site visit with local experts. The National Park Service held a GRI scoping meeting for Apostle Islands National Lakeshore on July 20-21, 2010 both out in the field on a boating site visit from Bayfield, Wisconsin, and at the headquarters building for the Great Lakes Network in Ashland, Wisconsin. Jim Chappell (Colorado State University [CSU]) facilitated the discussion of map coverage and Bruce Heise (NPS-GRD) led the discussion regarding geologic processes and features at the park. Dick Ojakangas from the University of Minnesota at Duluth and Laurel Woodruff from the U.S. Geological Survey presented brief geologic overviews of the park and surrounding area. Participants at the meeting included NPS staff from the park and Geologic Resources Division; geologists from the University of Minnesota at Duluth, Wisconsin Geological and Natural History Survey, and U.S. Geological Survey; and cooperators from Colorado State University (see table 2). -

Apostle Islands National Seashore
Apostle Islands National Seashore David Speer & Phillip Larson October 2nd Fieldtrip Report Table of Contents Introduction 1 Stop 1: Apostle Island Boat Cruise 1 Stop 2: Coastal Geomorphology 5 Stop 3: Apostle Islands National Seashore Headquarters 12 Stop 4: Northern Great Lakes Visitors Center 13 Stop 5: Montreal River 16 Stop 6: Conclusion 17 Bibliography 18 Introduction: On October 2nd, 2006, I and the other Geomorphology students took a field excursion to the Apostle Islands and the south shore of Lake Superior. On this trip we made five stops along our journey where we noted various geomorphic features and landscapes of the Lake Superior region. We reached the south coast of Lake Superior in Bayfield, Wisconsin at around 10:00am. Here we made our first stop which included the entire boat trip we took which navigated through the various islands in the Apostle Island National Lakeshore. Our second stop was a beach on the shore of Lake Superior where we noted active anthropogenic and environmental changes to a shoreline. Stop three was at the visitor center for the Apostle Island National Lakeshore where we heard a short talk by the resident park ranger as he explained some of the unique features of the islands and why they should be protected. After that we proceeded to our fourth stop which was at the Northern Great Lakes Visitor Center. Here we learned, through various displays and videos, how the region had been impacted by various events over time. Finally our fifth stop was at the point where the Montreal River emptied into Lake Superior (Figure 1). -

The Archeological Investigation of Four Lighthouse Complexes at the Western End of Lake Superior: the 1988 Testing Program Withi~Postle Islands National Lakeshore
()-/~'7 The Archeological Investigation of Four Lighthouse Complexes at the Western End of lake Superior: The 1988 Testing Program Withi~postle Islands National lakeshore National Park Service Midwest Archeological Center Ie PLEAS: I\ZTurm TO: TECHN1Cfil 1~!r-0:1MAT!O;l c::: :-:-:.::1 DENVC1 S:::;-:"":::: c;:::--;::1 ON M1CROF!Lf,l B&WScans r~i~TIO;~f~L f;~~:' ~Z~'J:C2 z .J'-t~ZO~5 -- --~~~------- ----~---- b - ----- ------------ THE ARCHEOLOGICAL INVESTIGATION OF FOUR LIGHTHOUSE COMPLEXES AT THE WESTERN END OF LAKE SUPERIOR: THE 1988 TESTING PROGRAM WITHIN APOSTLE ISLANDS NATIONAL LAKESHORE By Vergil E. Noble Midwest Archeological Center Technical Report No.8 United States Department of the Interior National Park Service Midwest Archeological Center Lincoln, Nebraska 1993 Ie ------ -- -- - -- --_.- - ----------------------------- --- ----- ABSTRACT During June and July of 1988, archeologists from the Midwest Archeological Center performed limited testing at four historic lighthouse complexes within Apostle Islands National Lakeshore. The lighthouses, which still protect shipping at the western end of Lake Superior, were scheduled for exterior restoration work. Specifically, immediate plans called for the installation of drainage systems about certain structures on Sand Island, Michigan Island, Outer Island, and Devils Island to mitigate continuing ground water damage to their foundations. Archeological investigations sought to assess the potential impacts to cultural resources in those areas of the light stations that would be disturbed by proposed developments. Shovel probes and controlled test excavations were used to examine the proposed drain alignments, as well as construction staging areas. No significant cultural resources were discovered during the five-week project that would warrant modification of the drainage systems or further archeological excavation prior to construction of these improvements. -

U.S. Coast Guard Historian's Office
U.S. Coast Guard Historian’s Office Preserving Our History For Future Generations Historic Light Station Information WISCONSIN ALGOMA PIERHEAD LIGHT (FRONT & REAR) Location: MOUTH OF THE AHNAPEE RIVER ON LAKE MICHIGAN, WISCONSIN Station Established: 1893 Year Current Tower(s) First Lit: 1932 Operational? YES Automated? YES 1973 Deactivated: N/A Foundation Materials: PIER Construction Materials: STEEL Tower Shape: CYLINDRICAL Markings/Pattern: RED Relationship to Other Structure: INTEGRAL Original Lens: FRESNEL Range: 16 Miles Characteristic: Red, Isophase 6 sec. HISTORICAL INFORMATION: Ahnapee grew rapidly in the mid to late 1870s and became the home to the largest commercial fishing fleet on Lake Michigan. When the government would not approve a lighthouse to mark the entrance to the harbor, locals erected a couple of post lights at the outer end of the two piers at the harbor entrance. Congress finally appropriated money to build some range lights in 1891 to mark the entrance of the harbor at Ahnapee which was renamed Algoma. The light was built in 1892 after new piers were completed but the light was not lit until the beginning of the 1893 shipping season. The piers were unique in that they were split and off-set. There was not a single continuous pier. To get to the lighthouse a bridge had to be built to span the gap between the off-set portions. The range initially consisted of a front post light and a wooden skeletal tower rear range. In 1895 the rear range tower had an upgrade to the lens. A fifth order lens was installed that increased the range from 9 miles to 11 miles. -
Wisconsin's John Muir
Wisconsin’s John Muir An Exhibit Celebrating the Centennial of the National Park Service “Oh, that glorious Wisconsin wilderness! “Everything new and pure in the very prime of the spring when Nature’s pulses were beating highest and mysteriously keeping time with our own!” “Wilderness is a necessity... Mountain parks and reservations are useful not only as fountains of timber and irrigating rivers, but as fountains of life.” This exhibit was made possible through generous support from the estate of John Peters and the Follett Charitable Trust Muir in Wisconsin “When we first saw Fountain Lake Meadow, on a sultry evening, sprinkled with millions of lightning- bugs throbbing with light, the effect was so strange and beautiful that it seemed far too marvelous to be real.” John Muir (1838–1914) was one of America’s most important environmental thinkers and activists. He came to Wisconsin as a boy, grew up near Portage, and attended the University of Wisconsin. After decades of wandering in the mountains of California, he led the movement for national parks and helped create the Sierra Club. But for much of his life, Muir’s call to protect wild places fell on deaf ears. Muir studied science in Madison but quit in 1863 without a degree, “...leaving one University for another, the Wisconsin University for the University of the Wilderness.” Muir’s letter to the classmate who taught him botany at UW The Movement for National Parks Yosemite Valley “Everybody needs beauty as well as bread, places to play in and pray in, where Nature may heal and cheer and give strength to body and soul alike.” In 1872, Congress named Yellowstone the first national park. -

Forest Bird Communities in the Apostle Islands of Wisconsin
FOREST BIRD COMMUNITIES IN THE APOSTLE ISLANDS OF WISCONSIN BY EDWARDBEALS N the past few years more ecologists have become aware that the description I of plant and animal communities as discrete ecological units is an over- simplification of community relationships. Among botanists, Gleason (1926) has proposed that, inasmuch as each plant species has environmental require- ments different from those of every other species, the classification of plant communities into separate and distinct associations is inadequate. Following this individualistic concept, many phytosociologists have developed vegeta- tional gradients based either on measurement of environmental factors (e.g., Ramensky, 1930; Whittaker, 1956) or on a direct analysis of the vegetation itself (e.g., Curtis and McIntosh, 1951; Brown and Curtis, 1952; Goodall, 1954; Horikawa and Okutomi, 1955; Bray and Curtis, 1957). Animal ecolo- gists have also used community gradients in studies of insects (Whittaker, 1952; Kato et al., 1955), copepods (Whittaker and Fairbanks, 1958), and birds (Bond, 1957). Because different bird species seldom if ever coincide in their ecological distributions, no discrete communities can be clearly defined except where there are sharp changes of environment. Therefore the present paper describes the bird populations within the forests of the Apostle Islands in Lake Superior in terms of a community gradient, relating these bird communities to their environmental framework. Scientific names of plants follow Gleason (1952). ACKNOWLEDGMENTS I wish to acknowledgewith gratitude the financial aid received from the Frank M. Chapman Memorial Fund, with which the project was completed. Part of the field work was accomplished while I was working on a vegetational survey of the islands for the Wis- consin Conservation Department, and the cooperation from the Department at the Bay field station is appreciated. -

Lighthouses – Clippings
GREAT LAKES MARINE COLLECTION MILWAUKEE PUBLIC LIBRARY/WISCONSIN MARINE HISTORICAL SOCIETY MARINE SUBJECT FILES LIGHTHOUSE CLIPPINGS Current as of November 7, 2018 LIGHTHOUSE NAME – STATE - LAKE – FILE LOCATION Algoma Pierhead Light – Wisconsin – Lake Michigan - Algoma Alpena Light – Michigan – Lake Huron - Alpena Apostle Islands Lights – Wisconsin – Lake Superior - Apostle Islands Ashland Harbor Breakwater Light – Wisconsin – Lake Superior - Ashland Ashtabula Harbor Light – Ohio – Lake Erie - Ashtabula Badgeley Island – Ontario – Georgian Bay, Lake Huron – Badgeley Island Bailey’s Harbor Light – Wisconsin – Lake Michigan – Bailey’s Harbor, Door County Bailey’s Harbor Range Lights – Wisconsin – Lake Michigan – Bailey’s Harbor, Door County Bala Light – Ontario – Lake Muskoka – Muskoka Lakes Bar Point Shoal Light – Michigan – Lake Erie – Detroit River Baraga (Escanaba) (Sand Point) Light – Michigan – Lake Michigan – Sand Point Barber’s Point Light (Old) – New York – Lake Champlain – Barber’s Point Barcelona Light – New York – Lake Erie – Barcelona Lighthouse Battle Island Lightstation – Ontario – Lake Superior – Battle Island Light Beaver Head Light – Michigan – Lake Michigan – Beaver Island Beaver Island Harbor Light – Michigan – Lake Michigan – St. James (Beaver Island Harbor) Belle Isle Lighthouse – Michigan – Lake St. Clair – Belle Isle Bellevue Park Old Range Light – Michigan/Ontario – St. Mary’s River – Bellevue Park Bete Grise Light – Michigan – Lake Superior – Mendota (Bete Grise) Bete Grise Bay Light – Michigan – Lake Superior -

Double-Crested Cormorant Damage to a Commercial Fishery in the Apostle Islands, Wisconsin
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln 2 - Second Eastern Wildlife Damage Control Conference (1985) Eastern Wildlife Damage Control Conferences September 1985 DOUBLE-CRESTED CORMORANT DAMAGE TO A COMMERCIAL FISHERY IN THE APOSTLE ISLANDS, WISCONSIN Scott R. Craven University of Wisconsin-Madison Esther Lev University of Wisconsin-Madison Follow this and additional works at: https://digitalcommons.unl.edu/ewdcc2 Part of the Environmental Health and Protection Commons Craven, Scott R. and Lev, Esther, "DOUBLE-CRESTED CORMORANT DAMAGE TO A COMMERCIAL FISHERY IN THE APOSTLE ISLANDS, WISCONSIN" (1985). 2 - Second Eastern Wildlife Damage Control Conference (1985). 11. https://digitalcommons.unl.edu/ewdcc2/11 This Article is brought to you for free and open access by the Eastern Wildlife Damage Control Conferences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in 2 - Second Eastern Wildlife Damage Control Conference (1985) by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. DOUBLE-CRESTED CORMORANT DAMAGE TO A COMMERCIAL FISHERY IN THE APOSTLE ISLANDS, WISCONSIN by Scott R. Craven and Esther Lev ABSTRACT eliminated perching and an old-fash- ioned scare-crow. National Park Service The endangered classification of policy precludes direct control of the double-crested cormorant (DCC) the increasing DCC population. in Wisconsin resulted in complete protection and significant management efforts in the 1970's., These efforts, INTRODUCTION probably coupled with reduced pesticide Double-crested cormorants loads, resulted in a resurgence of (Phalacrocorax auritus) have conflicted Wisconsin cormorant populations from with commercial fisheries along the a low of 66 pairs in 1972 to 1028 coast of Maine, in Wisconsin, in the pairs in 1982» The DCC was reclassified Great Lakes region in general (Matteson as a threatened species in 1982. -

Ashland County
APOSTLE ISLANDS AREA “Out & Back” & “Loop” Ashland County Bicycling Road Routes Ashland County Tourism Ashland Chamber of Commerce This bicycle map is designed to assist bicyclists in Bayfield & Ashland Counties & Visitor Bureau identifying enjoyable bicycling routes in Ashland & Bayfield Counties. 1716 Lake Shore Dr W • Ashland, WI 54806 715-682-2500 • 800-284-9484 These bicycle routes are intended for use by adult bicyclists who have WISCONSIN www.travelashlandcounty.com had driver’s training, and possess the skills and knowledge to handle www.visitashland.com shared bicycle/motor vehicle situations. Keep in mind that roadway Madeline Island Chamber of Commerce conditions may vary considerably by the day of the week, time of 715-747-2801 • 888-475-3386 day, weather conditions and construction. All bicyclists are encouraged www.madelineisland.com Sand Little Sand Bay Raspberry to select routes with riding conditions that match their own skill level Bay Mellen Area Chamber of Commerce Bay Rd. Bay and preparation. Apostle Islands 715-274-2330 • www.mellenwi.com National Lakeshore bears full responsibility for his or County Hwy K Old County Hwy K Peterson Hill The user of this map 13 Old Bayfield County her safety. Bicyclists should exercise good judgement and obey traffic Bark Pt. Siskiwit 10 Bay Bay Bayfield County Tourism laws on all roadways, regardless of the bicycle routes on this map. Rd. Red Cliff 117 E 5th St/PO Box 832 Happy Washburn, WI 54891 The state, Ashland and Bayfield Counties, cities, villages and towns, Compton and the agencies who have contributed to the development of this Cornucopia Robbs Rd. -

Amazing Apostle Islands Happy Times Tours & Experiences
Amazing Apostle Islands (Bayfield, WI) Trip Includes: - 3 Nights at the Legendary Waters Resort & Casino with a total of $75 in Legendary Loot (Promotional Play) - Drive through Scenic Northern Wisconsin and enjoy a shopping stop in Minocqua - Visit the Northern Great Lakes Visitor Apostle Islands – Sea Cave Raspberry Island Lighthouse Center - Ferry to Madeline Island Escape from the world for a while. Reconect with - Locally Guided Madeline Island Tour nature and enjoy a vacation you will never forget! - Lunch at the Pub on Madeline Island - Lighthouse & Sea Caves Cruise 4 Days: September 7 – 10, 2021 - Old Rittenhouse Inn Victorian Luncheon Day 1 B, D - Bayfield Guided Tour with a stop at a Enjoy a boxed breakfast as we head North. We will make a shopping stop in scenic Local Farm Minocqua, Wisconsin before continuing our journey through beautiful Northern - Visit Bonnie & Clyde’s Gangster Park Wisconsin. We will make a stop at the Northern Great Lakes Visitor Center to learn - Motorcoach Transportation and the a little more about this area before an included dinner is included at a local place known for their Prime Rib and Whitefish. We will arrive at the Legendary Waters Services of a Happy Times Tour Director Resort & Casino for a 3 night stay and all rooms have a beautiful view. Everyone - 7 Meals will receive a total of $75 in Legendary Loot (promotional play) during your stay at Departure Times & Locations: the Legendary Waters Resort & Casino. 6:30am Depart College Ave NE P&R (estimated return at 7:30pm) Day 2 B, L 7:00am Depart Watertown Plank P&R Today, we will board the Madeline Island Ferry to Madeline Island, where the (estimated return at 7:00pm) sandy shores and magnificent lake air beckons.