C. R,Darwin and J. D.Hooker
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CHAPTER 2 REVIEW of the LITERATURE 2.1 Taxa And
CHAPTER 2 REVIEW OF THE LITERATURE 2.1 Taxa and Classification of Acalypha indica Linn., Bridelia retusa (L.) A. Juss. and Cleidion javanicum BL. 2.11 Taxa and Classification of Acalypha indica Linn. Kingdom : Plantae Division : Magnoliophyta Class : Magnoliopsida Order : Euphorbiales Family : Euphorbiaceae Subfamily : Acalyphoideae Genus : Acalypha Species : Acalypha indica Linn. (Saha and Ahmed, 2011) Plant Synonyms: Acalypha ciliata Wall., A. canescens Wall., A. spicata Forsk. (35) Common names: Brennkraut (German), alcalifa (Brazil) and Ricinela (Spanish) (36). 9 2.12 Taxa and Classification of Bridelia retusa (L.) A. Juss. Kingdom : Plantae Division : Magnoliophyta Class : Magnoliopsida Order : Malpighiales Family : Euphorbiaceae Genus : Bridelia Species : Bridelia retusa (L.) A. Juss. Plant Synonyms: Bridelia airy-shawii Li. Common names: Ekdania (37,38). 2.13 Taxa and Classification of Cleidion javanicum BL. Kingdom : Plantae Subkingdom : Tracheobionta Superdivision : Spermatophyta Division : Magnoliophyta Class : Magnoliopsida Subclass : Magnoliopsida Order : Malpighiales Family : Euphorbiaceae Genus : Cleidion Species : Cleidion javanicum BL. Plant Synonyms: Acalypha spiciflora Burm. f. , Lasiostylis salicifolia Presl. Cleidion spiciflorum (Burm.f.) Merr. Common names: Malayalam and Yellari (39). 10 2.2 Review of chemical composition and bioactivities of Acalypha indica Linn., Bridelia retusa (L.) A. Juss. and Cleidion javanicum BL. 2.2.1 Review of chemical composition and bioactivities of Acalypha indica Linn. Acalypha indica -

2000 HSS/PSA Program 1
HISTORY OF SCIENCE SOCIETY 2000 ANNUAL MEETING PHILOSOPHY OF SCIENCE ASSOCIATION 2000 BIANNUAL MEETING 2-5 November 2000 Vancouver, British Columbia, Canada Hyatt Regency Vancouver CONTENTS Acknowledgments 3 HSS Officers, Program Chairs, and Council 4 PSA Officers and Program Committee 5 General Information 6 HSS / PSA 2000 Annual Meeting Book Exhibit Layout 7 Floor Plans: Hyatt Regency Vancouver 8-9 Vancouver Points of Interest 10-13 Committees and Interest Groups 14-15 HSS Full Program Schedule 16-20 HSS 2000 Program 21-43 HSS Distinguished Lecture 40 HSS Abstracts 44-187 PSA Full Program Schedule 188-190 PSA 2000 Program 191-202 PSA President’s Address 197 PSA Abstracts 203-245 HSS/PSA Program Index 246-252 Advertisements 253 Cover Illustration: SeaBus riders get the best view of Vancouver from the water. Offering regular service on the busiest routes from 5 a.m. to 2 a.m. and late night owl service on some downtown suburban routes until 4:20 a.m., Greater Vancouver’s transit system--the bus, SkyTrain and SeaBus-- covers more than 1800 square kilometers (695 square miles) of the Lower Mainland. The SkyTrain, a completely automated light rapid transit system, offers direct, efficient service between downtown Vancouver and suburban environs. It follows a scenic elevated 29 kilometer (18 mile) route with 20 stations along the way. All the SkyTrain stations, except Granville, have elevators and each train is wheelchair accessible. The SkyTrain links with buses at most of the 20 stations and connects with the SeaBus in downtown Vancouver. It operates daily, every two to five minutes. -

I.—Eminent Living Geologists : William Thomas Blanford, C.I.E., Ll.T)., F.E.S., V.P
THE GEOLOGICAL MAGAZINE. NEW SERIES. DECADE V. VOL. II. No. I —JANUARY, 1905. OBIGIITAL AETICLES. I.—EMINENT LIVING GEOLOGISTS : WILLIAM THOMAS BLANFORD, C.I.E., LL.T)., F.E.S., V.P. Zool. Soc, Treas. Geol. Soc. (WITH A PORTRAIT, PLATE I.) HAT India has been in the past 300 years to our Army as W a nursery in which our soldiers have obtained experience in their profession and earned their promotion, often to the highest rank, such in a lesser degree has it been to many of our geologists, who have, in the past much shorter period of 50 or 60 years, entered the service in this vast field of scientific enterprise, and, aided by a very few amateur geologists in the Army and of civilians attached to other branches of Government employ, have covered many thousand square miles of our Indian Empire with records of their untiring energy in the geological field. Among the amateurs may be recorded the names of Generals Sir Kichard Strachey and Sir Proby T. Cautley, Dr. Hugh Falconer, Lieut-Gen. C. A. McMahon; and as professional geologists, Dr. T. Oldham, H. B. Medlicott, J. G. Medlicott, Dr. Wm. King, Dr. Valentine Ball, the two Blanfords, W. Theobald, E. Bruce Foote, A. B. Wynne, C. L. Griesbach, E. D. Oldham, F. E. Mallet, C. S. Middlemiss, T. D. La Touche, Dr. F. Stoliczka, Professor W. Waagen, the present Director (T. H. Holland), and many others. Prominent among the earlier geological workers stand out the names of the brothers W. T. and H. F. Blanford, who joined the Indian Survey together in 1855. -
![Entry for ACALYPHA Acrogyna Pax [Family EUPHORBIACEAE]](https://docslib.b-cdn.net/cover/8673/entry-for-acalypha-acrogyna-pax-family-euphorbiaceae-78673.webp)
Entry for ACALYPHA Acrogyna Pax [Family EUPHORBIACEAE]
Entry for ACALYPHA acrogyna Pax [family EUPHORBIACEAE] http://plants.jstor.org/flora/flota011327 http://www.jstor.org Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the contributing partner regarding any further use of this work. Partner contact information may be obtained at http://plants.jstor.org/page/about/plants/PlantsProject.jsp. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Page 1 of 2 Entry for ACALYPHA acrogyna Pax [family EUPHORBIACEAE] Herbarium Royal Botanic Gardens, Kew (K) Collection Flora of Tropical Africa Resource Type Reference Sources Entry from Flora of Tropical Africa, Vol 6 Part 1, page 441 (1913) Author: (By J. G. Baker, with additions by C. H. Wright.) Names ACALYPHA acrogyna Pax [family EUPHORBIACEAE], in Engl. Jahrb. -

Charles Alfred Ernest Sheaf
Sir John Struthers PRCSEd 1895-7 (includes printed articles by his son, John William Struthers PRCSEd 1941-3) Reference and contact details: GB779 RCSEd GD/4 Location: RS R3 Title: Sir John Struthers Dates of Creation: 1839-1950 Held at: The Royal College of Surgeons of Edinburgh Extent: 460 items Name of Creator: Sir John Struthers (& J W Struthers) Language of Material: English. Level of Description: item Administrative/Biographical History: John Struthers (1823-1899) was born in Dunfermline, the second of three brothers, all of whom entered the medical profession. He took up surgery and was appointed to the Royal Infirmary following graduation. After a few years he abandoned surgery in favour of anatomy and was in time appointed Professor of Anatomy at Aberdeen. He held this post for 26 years, during which time he reorganised the whole teaching of anatomy and created an excellent museum of anatomy. After retiring from his Chair in Aberdeen he returned to Edinburgh where he devoted his energies to the Royal College of Surgeons. He was President from 1895 to 1897 and was knighted in 1898. In 1899 the Struthers Lecture was inaugurated and was given every third year until 1974. His son, John William Struthers M.D., LL.D. FRCSEd was President of the College from 1941-1943 UNIVERSITY NOTICES, 1839-59 GD4/1 1839, April 22nd Printed copies (4) of Bill introducing increased lecture fees in the department of Anatomy at the Edinburgh Medical School for the winter season, 1839-40. [4 sheets] GD4/2 1842, April Copy of the address given to students by Richard Huie, PRCSEd (1840-42), at the prize giving of the Edinburgh Medical School. -
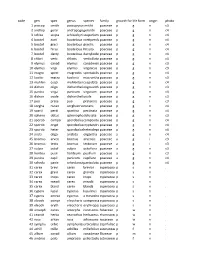
Species List (PDF)
code gen spec genus species family growth formlife form origin photo 1 pascop smith pascopyrumsmithii poaceae p g n c3 2 androp gerar andropogongerardii poaceae p g n c4 3 schiza scopa schizachyriumscoparium poaceae p g n c4 4 boutel curti bouteloua curtipendulapoaceae p g n c4 5 boutel graci bouteloua gracilis poaceae p g n c4 6 boutel hirsu bouteloua hirsuta poaceae p g n c4 7 boutel dacty bouteloua dactyloidespoaceae p g n c4 8 chlori verti chloris verticillata poaceae p g n c4 9 elymus canad elymus canadensispoaceae p g n c3 10 elymus virgi elymus virginicus poaceae p g n c3 11 eragro spect eragrostis spectabilis poaceae p g n c4 12 koeler macra koeleria macrantha poaceae p g n c3 13 muhlen cuspi muhlenbergiacuspidata poaceae p g n c4 14 dichan oligo dichantheliumoligosanthespoaceae p g n c3 15 panicu virga panicum virgatum poaceae p g n c4 16 dichan ovale dichantheliumovale poaceae p g n c3 17 poa prate poa pratensis poaceae p g i c3 18 sorgha nutan sorghastrumnutans poaceae p g n c4 19 sparti pecti spartina pectinata poaceae p g n c4 20 spheno obtus sphenopholisobtusata poaceae p g n c3 21 sporob compo sporoboluscomposituspoaceae p g n c4 22 sporob crypt sporoboluscryptandruspoaceae p g n c4 23 sporob heter sporobolusheterolepispoaceae p g n c4 24 aristi oliga aristida oligantha poaceae a g n c4 25 bromus arven bromus arvensis poaceae a g i c3 26 bromus tecto bromus tectorum poaceae a g i c3 27 vulpia octof vulpia octoflora poaceae a g n c3 28 hordeu pusil hordeum pusillum poaceae a g n c3 29 panicu capil panicum capillare poaceae a g n c4 30 schedo panic schedonnarduspaniculatuspoaceae p g n c4 31 carex brevi carex brevior cyperaceaep s n . -

Leaf Epidermal Studies of Three Species of Acalypha Linn
Available online a t www.pelagiaresearchlibrary.com Pelagia Research Library Advances in Applied Science Research, 2012, 3 (5):3185-3199 ISSN: 0976-8610 CODEN (USA): AASRFC Leaf epidermal studies of three species of Acalypha Linn. (Euphorbiaceae) 1* Essiett Uduak Aniesua and 1Etukudo Inyene Silas Department of Botany and Ecological Studies, University of Uyo, Uyo _____________________________________________________________________________________________ ABSTRACT Leaf epidermal studies of three species of Acalypha are described. The mature stomata were laterocytic, staurocytic, anisocytic, paracytic and diacytic. The abnormalities noticed include unopen stomatal pore, two stomata sharing one subsidiary cell, one guard cell, parallel contiguous and aborted guard cell. A. godseffiana can be distinguished by parallel contiguous on both surfaces. Curved uniseriate non-glandular trichomes were restricted to A. wilkesiana. Two stomata sharing one subsidiary cell occurred only on the lower surface of A. hispida. The shapes of epidermal anticlinal cell walls, guard cell areas, stomata index and trichomes varied. The differences are of taxonomic importance and can be used to identify and delimit each species by supporting other systematic lines of evidence. Keywords: Acalypha species, Epidermal, Stomata, Nigeria, Euphorbiaceae. _____________________________________________________________________________________________ INTRODUCTION Euphorbiaceae, the spurge family of the flowering plants with 500 genera and around 7,500 species. Most are herbs, -

Last Name, First Name ECOL 249. Quiz 5 Part I. Answer Twelve
May not be posted online without written permission of W. M. Schaffer, Univ. AZ., Tucson, AZ. ___________________________ Last Name, First Name ECOL 249. Quiz 5 Part I. Answer twelve (12) of the following questions (5 points each). Only the first 12 answers will be graded. 1. By 1872, Vestiges had a. been banned for atheistic and seditious content. b. been embraced by the physicists who approved its endorsement of the nebula hypothesis. c. been outsold 5:1 by The Origin d. outsold The Origin by about 2:1 [Lecture V, 3] e. had so enraged the public that its formerly anonymous author was forced to flee the country with wife and children. 2. Which of the following scientific ideas was not endorsed by Vestiges? a. Nebular hypothesis. b. Progress in the fossil record. c. Quinerian classification. d. Spontaneous generation of mites e. wants and ... exercise... in the way suggested by Lamarck. [Lecture V, 9] 3. The ideas of _________ were eventually confirmed by the discovery of dorso-ventral patterning inversion in chordates and invertebrates. a. É. Geoffroy St. Hilaire [Lecture V, 62-63] b. Ernst Haeckel c. K. E. von Baer d. Robert Grant e. Richard Owen 4. According to Adrian Desmond (Designing the Dinosaur), Owen’s creation of the order Dinosauria and his mammal-like dinosaur reconstructions were motivated by antipa- thy to ___________ . a. Edward Forbes b. Louis Agassiz Megaloceros, a bipedal theropod dinosaur, as im- c. Robert Chambers. agined by Owen and restored by Waterhouse d. Robert Grant [Desmond, 1987, 224 ff] Hawkins for the Crystal Palace Exhibition. -

''It Ain't Over 'Til It's Over'': Rethinking the Darwinian Revolution
Journal of the History of Biology (2005) 38: 33–49 Ó Springer 2005 DOI 10.1007/s10739-004-6508-z ‘‘It Ain’t Over ‘til it’s Over’’: Rethinking the Darwinian Revolution VASSILIKI BETTY SMOCOVITIS Department of Philosophy and History of Science University of Athens, Athens Greece and Departments of Zoology and History University of Florida Gainesville, FL 32611 USA E-mail: [email protected]fl.edu Abstract. This paper attempts a critical examination of scholarly understanding of the historical event referred to as ‘‘the Darwinian Revolution.’’ In particular, it concentrates on some of the major scholarly works that have appeared since the publication in 1979 of Michael Ruse’s The Darwinian Revolution: Nature Red in Tooth and Claw. The paper closes by arguing that fruitful critical perspectives on what counts as this event can be gained by locating it in a range of historiographic and disciplinary contexts that include the emergence of the discipline of evolutionary biology (following the ‘‘evolutionary synthesis’’), the 1959 Darwin centenary, and the maturation of the discipline of the history of science. Broader perspectives on something called the ‘‘Darwinian Revolu- tion’’ are called for that include recognizing that it does not map a one-to-one corre- spondence with the history of evolution, broadly construed. Keywords: Darwin centennial, Darwinian Revolution, discipline, evolutionary biology evolutionary synthesis, historiography The Darwinian Revolution was probably the most significant revolution that has ever occurred in the sciences, -

Natural Selection: Charles Darwin & Alfred Russel Wallace
Search | Glossary | Home << previous | next > > Natural Selection: Charles Darwin & Alfred Russel Wallace The genius of Darwin (left), the way in which he suddenly turned all of biology upside down in 1859 with the publication of the Origin of Species , can sometimes give the misleading impression that the theory of evolution sprang from his forehead fully formed without any precedent in scientific history. But as earlier chapters in this history have shown, the raw material for Darwin's theory had been known for decades. Geologists and paleontologists had made a compelling case that life had been on Earth for a long time, that it had changed over that time, and that many species had become extinct. At the same time, embryologists and other naturalists studying living animals in the early 1800s had discovered, sometimes unwittingly, much of the A visit to the Galapagos Islands in 1835 helped Darwin best evidence for Darwin's formulate his ideas on natural selection. He found theory. several species of finch adapted to different environmental niches. The finches also differed in beak shape, food source, and how food was captured. Pre-Darwinian ideas about evolution It was Darwin's genius both to show how all this evidence favored the evolution of species from a common ancestor and to offer a plausible mechanism by which life might evolve. Lamarck and others had promoted evolutionary theories, but in order to explain just how life changed, they depended on speculation. Typically, they claimed that evolution was guided by some long-term trend. Lamarck, for example, thought that life strove over time to rise from simple single-celled forms to complex ones. -

The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886
The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 Sascha Nolden, Simon Nathan & Esme Mildenhall Geoscience Society of New Zealand miscellaneous publication 133H November 2013 Published by the Geoscience Society of New Zealand Inc, 2013 Information on the Society and its publications is given at www.gsnz.org.nz © Copyright Simon Nathan & Sascha Nolden, 2013 Geoscience Society of New Zealand miscellaneous publication 133H ISBN 978-1-877480-29-4 ISSN 2230-4495 (Online) ISSN 2230-4487 (Print) We gratefully acknowledge financial assistance from the Brian Mason Scientific and Technical Trust which has provided financial support for this project. This document is available as a PDF file that can be downloaded from the Geoscience Society website at: http://www.gsnz.org.nz/information/misc-series-i-49.html Bibliographic Reference Nolden, S.; Nathan, S.; Mildenhall, E. 2013: The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886. Geoscience Society of New Zealand miscellaneous publication 133H. 219 pages. The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 CONTENTS Introduction 3 The Sumner Cave controversy Sources of the Haast-Hooker correspondence Transcription and presentation of the letters Acknowledgements References Calendar of Letters 8 Transcriptions of the Haast-Hooker letters 12 Appendix 1: Undated letter (fragment), ca 1867 208 Appendix 2: Obituary for Sir Julius von Haast 209 Appendix 3: Biographical register of names mentioned in the correspondence 213 Figures Figure 1: Photographs -

Former Fellows Biographical Index Part
Former Fellows of The Royal Society of Edinburgh 1783 – 2002 Biographical Index Part Two ISBN 0 902198 84 X Published July 2006 © The Royal Society of Edinburgh 22-26 George Street, Edinburgh, EH2 2PQ BIOGRAPHICAL INDEX OF FORMER FELLOWS OF THE ROYAL SOCIETY OF EDINBURGH 1783 – 2002 PART II K-Z C D Waterston and A Macmillan Shearer This is a print-out of the biographical index of over 4000 former Fellows of the Royal Society of Edinburgh as held on the Society’s computer system in October 2005. It lists former Fellows from the foundation of the Society in 1783 to October 2002. Most are deceased Fellows up to and including the list given in the RSE Directory 2003 (Session 2002-3) but some former Fellows who left the Society by resignation or were removed from the roll are still living. HISTORY OF THE PROJECT Information on the Fellowship has been kept by the Society in many ways – unpublished sources include Council and Committee Minutes, Card Indices, and correspondence; published sources such as Transactions, Proceedings, Year Books, Billets, Candidates Lists, etc. All have been examined by the compilers, who have found the Minutes, particularly Committee Minutes, to be of variable quality, and it is to be regretted that the Society’s holdings of published billets and candidates lists are incomplete. The late Professor Neil Campbell prepared from these sources a loose-leaf list of some 1500 Ordinary Fellows elected during the Society’s first hundred years. He listed name and forenames, title where applicable and national honours, profession or discipline, position held, some information on membership of the other societies, dates of birth, election to the Society and death or resignation from the Society and reference to a printed biography.