N,OXBK Entod60logtise FORENIMQ '
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

KOMMUNEPLAN for OSLO Status Og Videre Utvikling
KOMMUNEPLAN FOR OSLO status og videre utvikling grape architects KLIMA OG FORTETTING Bærekraftig vekst er svaret grape architects KOMMUNEPLAN FOR OSLO_status og fremtidig utvikling_06.03.11 2 grape architects KLIMA OG FORTETTING Atlanta vs Barcelona ATLANTA - 1225 innbyggere per km2 BARCELONA - 32 900 m2 per km2 fra The New Climate Economy_chapter 2_Cities: KOMMUNEPLAN FOR OSLO_status og fremtidig utvikling_06.03.11 3 grape architects HVA GIR EN GOD BY? DISCUSSION NOTE 3 UN HABITAT URBAN PLANNING A NEW STRATEGY OF SUSTAINABLE NEIGHBOURHOOD PLANNING: FIVE PRINCIPLES UN-Habitat supports countries to develop urban THE FIVE PRINCIPLES ARE: planning methods and systems to address current urbanization challenges such as population growth, 1. Adequate space for streets and an efficient street network. The street urban sprawl, poverty, inequality, pollution, network should occupy at least 30 per cent of congestion, as well as urban biodiversity, urban the land and at least 18 km of street length mobility and energy. per km². 2. High density. At least 15,000 people per In recent decades, the landscape of Cities of the future should build a km², that is 150 people/ha or 61 people/acre. cities has changed significantly because different type of urban structure and konnektivitet - tetthet - variasjon of rapid urban population growth. A space, where city life thrives and the 3. Mixed land-use. At least 40 per cent of floor major feature of fast growing cities most common problems of current space should be allocated for economic use in is urban sprawl, which drives the urbanization are addressed. UN-Habitat any neighbourhood. occupation of large areas of land and is proposes an approach that summarizes usually accompanied by many serious and refines existing sustainable urban 4. -
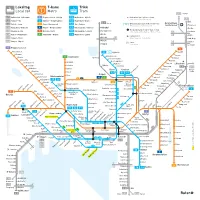
Lokaltog Local Rail Trikk Tram T-Bane Metro
Lokaltog T-bane Trikk Local rail Metro Tram L12 Eidsvoll L 1 Spikkestad – Lillestrøm 1 Frognerseteren – Helsfyr 11 Majorstuen – Kjelsås Holdeplass bare i pilens retning Stop in direction of arrow only L13 L 2 Skøyen – Ski 2 Gjønnes – Ellingsrudåsen 12 Majorstuen – Disen Dal L 3 Jaren Oslo lufthavn L 3 Oslo S – Jaren 3 Storo – Mortensrud 13 Jar – Grefsen 12 Endeholdeplass bare til bestemte tider Final stop at certain times only Gardermoen Hauerseter L12 Kongsberg – Eidsvoll 4 Ringen – Bergkrystallen 17 Rikshospitalet – Grefsen Hakadal Nordby Overgangsmuliget Tog / T-bane / Trikk Varingskollen L13 Drammen – Dal 5 Østerås – Vestli 18 Rikshospitalet – Holtet Interchange option Railway / Metro / Tram 4N Jessheim Åneby L14 Asker – Kongsvinger 6 Sognsvann – Ringen 19 Majorstuen – Ljabru Kløfta Flytogstasjon 3Ø Nittedal L21 Skøyen – Moss 2Ø Airport Express Train station Lindeberg Movatn 1 L22 Skøyen – Mysen Soner 3Ø Frogner Snippen 2V Fare zones 2Ø Leirsund 1 Frognerseteren 5 Voksenkollen 11 12 Kjelsås Vestli Lillevann Kjelsåsalleen Stovner Skogen 6 Sognsvann Kjelsås Grefsen stadion Rommen Voksenlia Grefsenplatået Romsås Kringsjå Holmenkollen Glads vei Grorud Lillestrøm Besserud Holstein Nydalen Sanatoriet Ammerud L 1 L14 Midtstuen Østhorn Disen Grefsen Kalbakken Sagdalen Kongs- Skådalen Tåsen Rødtvet vinger 12 13 17 Sinsenkrysset Strømmen Vettakollen Ringen Berg Veitvet Fjellhamar Gulleråsen Rikshospitalet Linderud 3 4 4 6 Hanaborg Gråkammen 17 18 Vollebekk Lørenskog Storo Sinsen Slemdal Nydalen 3 Risløkka Høybråten 2Ø Gaustad- Ullevål stadion -

Policies for Sustainable Commuting
TØI report 1527/2016 Farideh Ramjerdi Kåre H Skollerud Jørgen Aarhaug Policies for sustainable commuting TØI-rapport 1527/2016 Policies for sustainable commuting Farideh Ramjerdi, Kåre H. Skollerud, Jørgen Aarhaug Picture front page: Tidsskriftet Samferdsels temaarkiv This report is covered by the terms and conditions specified by the Norwegian Copyright Act. Contents of the report may be used for referencing or as a source of information. Quotations or references must be attributed to the Institute of Transport Economics (TØI) as the source with specific mention made to the author and report number. For other use, advance permission must be provided by TØI. ISSN 0808-1190 ISBN 978-82-480-1774-5 Electronic version Oslo, March 2017 Tittel Tiltak for bærekraftige arbeidsreiser Title Policies for sustainable commuting Forfatter(e): Farideh Ramjerdi, Kåre H. Author(s) Farideh Ramjerdi, Kåre H. Skollerud Jørgen Aarhaug Skollerud Jørgen Aarhaug Dato: 12.2016 Date: 12.2016 TØI-rapport 1527/2016 TØI Report: 1527/2016 Sider: 63 Pages: 63 ISBN elektronisk: 978-82-480-1774-5 ISBN Electronic: 978-82-480-1774-5 ISSN: 0808-1190 ISSN: 0808-1190 Finansieringskilde(r): Regionalt forskingsfond Financed by: The Regional Research Council hovedstaden, Akershus for Oslo and Akershus, Akershus fylkeskommune, Ruter AS, County, Ruter AS, The Jernbaneverket, Oslo kommune, Norwegian Railway Prosjekt: 3993 – Reisevaneendring i Oslo Project: 3993 – travel behavio ur change og Akershus – en analyse av in Oslo and Akershus – a study seks trafikk knutepunkt of six key -

TOUR GUIDE How to Find Oslo’S Rivers and Streams by Train, Tram, Bus and Metro OSLO RIVER FORUM
TOUR GUIDE How to find Oslo’s rivers and streams by train, tram, bus and Metro OSLO RIVER FORUM Oslo River Forum (Oslo Elveforum, OE) was founded in the year 2000 as a forum for volunteer groups and associations working for Oslo’s ten river catchments. Our task is to promote to the protection, rehabilitation and re-opening of the rivers and streams in the City’s built-up area. We work to make Oslo’s waterways clear, clean, open and accessible for citizens and visitors. Riversides are to be protected and turned into green space, with trails along each stream. Waters are to be healthy and aquatic species, including fish, encouraged. Oslo River Forum submits comments in all plan and zoning cases that concern the streams, protesting when develop- ments are proposed within the 20 and 12 metre limits given in the City’s zoning guidelines. We arrange guided tours along our rivers. And support schools, that adopt a part of their local stream. In a wet and wilder climate, we actively front the issue of re-opening the buried reaches of Oslo’s streams. Our vision: Living streams from forest to fjord. www.osloelveforum.no CONTENTS The terms ‘river ‘, ‘stream’, ‘brook’ or ‘creek’ each convey the size of a watercourse. In Oslo, three words apply: elv, bekk and å. A distinction is that one can leap over en bekk but not ei elv. From Danish there is a saying: Mange bekker små gjør en stor å. ’Many brooks make a large stream’, so å is larger than ‘bekk’. -

Vegetation Auf Den Schalenablagerungen in Bohuslän, Schweden
ACfA PHYTOGEOGRAPHICA SUECICA 56 EDIDIT SVENSKA VÄXTGEOGRAFISKA SÄLLSKAPET Vegetation · auf den Schalenablagerungen in Bohuslän, Schweden von H. Peter Hallberg Summary: Vegetation on Shell Deposits in Bohuslän, Sweden UPPSALA 1971 1 ALMQVIST & Wl�SELLS BOKTRYCKERI AB ACTA PHYTOGEOGRAPHICA SUECICA 56 Vegetation auf den Schalenablagerungen in Bohuslän, Schweden von H. Peter Hallberg Fil. lic., Göteborgs Nation Summary: Vegetation on Shell Deposits in Bohuslän, Sweden Akademische Abhandlung welche zur Erlangung der Doktorwürde mit Genehmigul!g der Mathematisch-Naturwissenschaftlichen Fakultät der Universität Uppsala am 19. März 1971, 10 Uhr vormittags, im Hörsaal des Pflanzenbiologischen Instituts öffentlich verteidigt wird UPPSALA 1971 Druckkostenbeitrag aus Längmanska Kulturfonden <0 H. Peter Hallberg 1971 Gedruckt in Schweden Almqvist & Wiksells Boktryckeri AB U ppsala 1971 VORWO RT Eines meiner grössten Interessen galt während meiner Bibliothekar A. Sjödin, bei dem ich meine ersten Schulzeit dem Sammeln und Pressen von Pflanzen. Erfahrungen in der Bibliotheksarbeit machte, was Diese Sammelleidenschaft führte mich frühzeitig mit mir später in anderem Zusammenhang zugutekam. zwei älteren Rotanisten zusammen, mit Doc. V. Gill Während meiner Reisen nach Bohuslän und ander ner und Lektor R. Ivarsson. Ich lernte beide während wärts traf ich mit vielen Menschen zusammen, die zahlreicher Exkursionen kennen - nicht zuletzt mit mir in manchem hilfreich beigestanden und mir dem Verein Natura an Hvitfeldtska läroverket in Gö jeweils ihre reichen Erfahrungen vermittelt haben. teborg während der vierziger Jahre -, die in Gillner Verschiedene Spezialisten haben bei der Bestim oft einen inspirierenden Leiter hatten. Da wir drei uns mung von schwer zu bestimmenden Pflanzen gehol später in unseren Forschungen an Växtbiologiska in fen oder meine Bestimmungen kontrolliert: bei Ta stitutionen mit der Vegetation Bohusläns befassten, er raxacum - Herr T. -

Urbact Case Hasle
URBACT CASE HASLE REINVENTING THE FRINGE FEASIBILITY STUDY Involved team Hans Baalerud / Karoline Birkeli-Gauss / Håkon Ellingsen Per Christian Stokke / Rune Skeie / Ida Tesaker Belland Karin Edlund / Hans Otte / Hoegh Eiendom / Femke Peters / Eirik Stokke 0. Summary Asplan Viak has been awarded the task of presenting a study on how to transform a brownfield site currently housing the national arenas for Tennis and Gymnastic in Oslo. These large halls occupy the space in an inefficient way as well as holding fairly poor architectural qualities. A local kindergarten is also on site. At the same time, the municipal landlord, EBY, wishes to develop the site as the area has grown attractive to investors and is undergoing huge transformations. However, the sport associations hold leasing contracts running until 2054. Therefore, should the site be developed, one should investigate how to include the existing programs whilst adding new functions. The project is a study on how to co-use and how to arrange the sport facilities in a more logical and urban way, giving space for other functions to revitalise the plot. And not least synergies that may occur. The project has designed two coherent designs that test density and quality in different ways, using a similar program throughout the site. By adding new functions, mostly housing, the project uses the added programs as a financial incentive to fund the reestablishment of the sports halls. Further, the project showcases innovative programming on how to design and facilitate for better co-use and alternative housing programs. These are more efficient designs compared to traditional urban planning, and follow the principles of a sharing and circular economy. -

Social Media and European Politics
Workshop: Social Media and European Politics Comfort Hotel Grand Central Jernbanetorget 1 Tel: +47 22 98 28 00 https://www.nordicchoicehot els.no/comfort/comfort-hotel- grand-central/ ARENA Centre for European Studies University of Oslo Sognsveien 68 The workshop is held at ARENA’s premises, some four kilometres north of Oslo’s city centre. You will find us on the 2nd floor in a red brick building opposite to the foot- ball stadium. The building bears a sign with 'Univer- sitetet i Oslo' and the entrance is be- tween the buildings of number 68 and 70. From the city centre to ARENA The metro station (T-bane) at the central train station Jernbanetorget is connected to the station Ullevål Stadion by the westbound lines #3 Storo, #4 Ringen or #6 Sognsvann (six stops). Jernbanetorget is the station very close to your hotel. From Nationaltheatret you can take the same lines, only four stops. All lines leave from the same platform. Tickets must be purchased in advance. A single ticket costs 30 NOK and can be pur- chased at ticketing machines at most metro stations, in most kiosks and using the ‘RuterBillett’ app (see more here: https://ruter.no/en/buying-tickets/tickets-and- fares/single-tickets/). Oslo Airport Gardermoen (OSL) Oslo Airport Gardermoen is roughly 50 km north of Oslo, and the Airport Express Train (Flytoget) is the fastest way of getting to the city centre. The train leaves every 10 minutes from Oslo Airport Gardermoen to Oslo Central Station (Oslo S), and eve- ry 20 minutes to the station Nationaltheatret (train continuing to Drammen). -
Skinne Mars 2015
Lokaltog T-bane Trikk Local rail Metro Tram L12 Eidsvoll L 1 Spikkestad – Lillestrøm 1 Frognerseteren – Helsfyr 11 Majorstuen – Kjelsås Holdeplass bare i pilens retning Stop in direction of arrow only L13 L 2 Skøyen – Ski 2 Kolsås – Ellingsrudåsen 12 Majorstuen – Disen (Kjelsås) Dal L 3 Oslo lufthavn L 3 Oslo S – Jaren 3 Sinsen – Mortensrud 13 Bekkestua – Grefsen Jaren 12 Endeholdeplass bare til bestemte tider Final stop at certain times only Gardermoen Hauerseter L12 Kongsberg – Eidsvoll 4 Ringen – Bergkrystallen 17 Rikshospitalet – Grefsen Hakadal Nordby Overgangsmuliget Tog / T-bane / Trikk Varingskollen L13 Drammen – Dal 5 Østerås – Vestli 18 Rikshospitalet – Holtet (Ljabru) Interchange option Railway / Metro / Tram 4N Jessheim Åneby L14 Asker – Kongsvinger 6 Sognsvann – Ringen 19 Majorstuen – Ljabru Kløfta Flytogstasjon 3Ø Nittedal L21 Skøyen – Moss 2Ø Airport Express Train station Lindeberg Movatn 1 L22 Skøyen – Mysen Soner 3Ø Frogner Snippen 2V Fare zones 2Ø Leirsund 1 Frognerseteren 5 Voksenkollen 11 12 Kjelsås Vestli Lillevann Kjelsåsalleen Stovner Skogen 6 Sognsvann Kjelsås Grefsen stadion Rommen Voksenlia Grefsenplatået Romsås Kringsjå Holmenkollen Glads vei Grorud Lillestrøm Holstein Nydalen Besserud Doktor Smiths vei Ammerud set L 1 L14 Midtstuen Østhorn Disen Grefsen Kalbakken Sagdalen Kongs- Skådalen Tåsen Rødtvet vinger 12 13 17 Sinsenkrys Strømmen Vettakollen Berg Veitvet Fjellhamar Gulleråsen Rikshospitalet Linderud Hanaborg Gråkammen 17 18 Vollebekk Storo Sinsen Lørenskog Slemdal Nydalen 4 6 Risløkka Gaustad- Ullevål -
University of Minnesota
THE UNIVERSITY OF MINNESOTA Announces Its );tareft eommcllecmcllt 1963 NORTHROP MEMORIAL AUDITORIUM SATURDAY EVENING, MARCH 23 AT EIGHT THIRTY O'CLOCK Universitv of Minnesota THE BOARD OF REGENTS* Dr. O. Meredith Wilson, President Mr. Laurence R. Lunden, Secretary Mr. Clinton T. Johnson, Treasurer Mr. Sterling B. Garrison, Assistant Secretary The Honorable Charles W. Mayo, M.D., Rochester First Vice President and Chairman The Honorable Marjorie J. Howard (Mrs. C. Edward), Excelsior Second Vice President The Honorable Daniel C. Gainey, Owatonna The Honorable Richard L. Griggs, Duluth The Honorable Bjarne E. Grottum, Jackson The Honorable Robert E. Hess, White Bear Lake The Honorable Fred J. Hughes, St. Cloud The Honorable A. I. Johnson, Benson The Honorable Lester A. Malkerson, Minneapolis The Honorable A. J. Olson, Renville The Honorable Otto A. Silha, Minneapolis The Honorable Herman F. Skyberg, Fisher *As of March 12, 1963. SMOKING AND USE OF CAMERAS AND RECORDERS-It is requested, by action of the Board of Regents, that in Northrop Memorial Auditorium smoking be confined to the outer lobby on the main floor, to the gallery lobbies, and to the lounge rooms. The use of cameras or tape recorders in the auditorium by members of the audio ence is prohibited. ?:ltis /s V(Jllr Univcrsitg CHARTERED in February, 1851, by the Legislative Assembly of the Territory of Minnesota, the University of Minnesota this year celebrated its one hundred and twelfth birthday. As one of the great Land-Grant universities in the nation, the University of Minnesota is dedicated to training the young people of today to become the leaders of tomorrow. -

Skilt for Syklister
Zinoberveien Bomveien Lille Aklungen Sorkedalsveien Store Gryta Lilloseterveien Stokkvann Rødkleivfaret Tryvannsveien Øvresetertjern NITTEDAL Strømsdammen Øvreseterveien Lillevann T LILLEVANN T VOKSENKOLLEN Gryteveien Kringlebekkveien Breisjøen T FROGNERSETEREN Setervollveien Voksenkollveien Kallandveien Haugakollveien Maridalsvannet Lysebuveien Ullveien FROGNERSETEREN Steinbruvann Sørkedalsveien Gamle Trondheimsvei Øvreseterveien Lillevannsveien Orreskogen Kringla Setertjern Svartkulp Ragnhild Schibbyes vei Voksenkollveien Sognsvann Jegersborgdammen Maridalsveien T SKOGEN Bankallstubben Blåbærsvingen T VESTLI Jerpefaret Trondheimsveien Hukenveien Nico Hambros vei Orrebakken Solemskogveien Thorleif Haugs vei Lachmanns vei Arnulf Øverlands vei Grindbakken n kollveie Orreveien Voksen Sognsvann Langevann Martha Tynes' vei Lillevannsveien Gjøkbakken Sverre Iversons vei Hospitsveien Brekkeveien Ytre Ringvei Langsetveien Ammerudveien Karen Platous vei Vestlisvingen Rundhaugveien Holmenkollveien Gjerdesmutten Svarttjern Bogstadvannet Hospitsveien Radioveien Fjellhøiveien Lachmanns vei T SOGNSVANN Frysjaveien Ammerudgrenda Inga Bjørnsons vei Sørkedalsveien Vettaliveien Voksenliveien Svarttrostveien Kjelsåsveien Alundamveien T VOKSENLIA Frysja RønningveienJetteveien Ellen Gleditsch vei Olaf Bulls vei Lilloseterveien Kongeveien Frognerseterveien AsbjørnsensKJELSÅS vei en Tokerudberget Ammerudg Margrethe Parms vei Olav M. Troviks vei Grinda Skjoldveien Midtoddveien Ravnkollbakken Røslyngveien Øvre LangåsØvre vei Skjoldvei Alnsjøen Parkenga Odvar -

NGT 33 3 4 235-250.Pdf
NORSK GEOLOGISK FORENING Møte 345 torsdag 5. mars 1953 på Farmasøytisk Institutt, Blindern. Tilstede 25 medlemmer og 7 gjester. Det ble meddelt at en vil søke å trykke foredragsreferat i tids skriftet. Spørsmålet om også å trykke diskusjonsinnlegg ble reist av Dietrichson. Det ble meddelt at styret hadde mottatt fra redaksjonen i Bergen bind 31 og bind 32 hefte l av tidsskriftet. Foredrag av R. C. Sørbye (Bergen): Kaledonidene i nord-østre Ryfylke og på H augesundshalvøya. På de eldre geologiske kart fra dette området, som stort sett dekkes av de topografiske gradteigkart Vikedal og Haugesund, finner vi tre atskilte bergartformasjoner: l. Grunnfjell, 2. Fyllittavdeling, 3. Høyfjellsgneis. Denne tre-inndeling kan for oversiktens skyld ennå brukes, hvis vi istedenfor <<grunnfjell)> skriver <<basalbergartene)>. Nyere undersøkelser på gradteig Haugesund viser nemlig en så kraftig påvirkning av den kaledonske orogenese i dette strøk, at sik r e pre-kambriske bergarter ikke lenger kan finnes vest for grensen Nedstrand- Bjoestrand, altså i det område som kalles Haugalandet. Min oppgave er å søke å påvise intensitetsgraden av den kale donske orogenese i de tre bergartformasjoner fra Suldalshei til Bøm lafjord. Forholdene langs grensene mellom disse formasjonene samt metamorfosen i fyllittene eller glimmerskifrene har hittil gitt de beste bidrag til løsning av oppgaven. I Suldalshei er grensen autokton, idet der er funnet udeformert basalkonglomerat, og de underliggende grunnfjellsbergartene er dis kordant mot grensen. De tilnærmet horisontale kaledonske skyv ninger har her foregått oppe i fyllittformasjonen, og grensen mellom fyllittavdelingen og høyfjellsgneisene er en tydelig skyvegrense. I området mellom Saudafjord - Sandsfjord og Haugalandets basalkompleks har skyvningene også gått for seg langs denne grensen, 6• - Geo!. -

Real Estate Projects
Real Estate Projects FEBRUARY 2018 Index Our vision and philosophy 4 All eyes on the occupier market 6 Storogården 8 Exited investments 10 Portal Skøyen 20 Present Investments 22 Pontoppidansgate 7, Oslo 24 Hauketoveien 8, Oslo 26 Industrigata 1, Lier 28 Vitaminveien 1, Oslo 30 Jessheim Næringspark, Jessheim 32 Beddingen 8, Oslo 34 Eikveien 19-20, Drammen 36 Trondheimsveien 113, Oslo 38 Drammensveien 211-213, Oslo 40 Hanaveien 17, Sandnes 42 Infarten 2, Kungsbacka 44 46 Our team 48 Contact us! Real Estate Projects February 2018 Liquidity and volume are also central to one’s ability to purchase and to reduce exit risk. Greater Oslo normally has over 60% of all transactions in the Norwegian market. By focusing on a limited geographical area our compet- itive advantage in all phases of the property life cycle is enhanced: identifying opportunities, understanding tenants locally, zoning, asset management, property development and exit strategy. Our vision is We have always focused on commuting hubs. In 2017 we bought two properties located at commuting hubs in “to transform Oslo, both within Oslo Municipality’s strategic area devel- opment plan towards 2030. This means that the potential bricks into gold” exists to increase the properties’ volume. Vitaminveien 1A at Storo was the largest acquisition last year at NOK 523 million and 21,000 m2. We have since closing in March rebranded the property with new signs and new façade paint, and approved building permit for The vision portrays who we are, an additional 2,300 m2 to transform the building from an our investment philosophy and the introverted brown brick block to an inviting modern of- results we wish to achieve.