Biodiversity: Implications for Monitoring, Science and Adaptive Planning 3 BIODIVERSITY
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Diseases of Trees in the Great Plains
United States Department of Agriculture Diseases of Trees in the Great Plains Forest Rocky Mountain General Technical Service Research Station Report RMRS-GTR-335 November 2016 Bergdahl, Aaron D.; Hill, Alison, tech. coords. 2016. Diseases of trees in the Great Plains. Gen. Tech. Rep. RMRS-GTR-335. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 229 p. Abstract Hosts, distribution, symptoms and signs, disease cycle, and management strategies are described for 84 hardwood and 32 conifer diseases in 56 chapters. Color illustrations are provided to aid in accurate diagnosis. A glossary of technical terms and indexes to hosts and pathogens also are included. Keywords: Tree diseases, forest pathology, Great Plains, forest and tree health, windbreaks. Cover photos by: James A. Walla (top left), Laurie J. Stepanek (top right), David Leatherman (middle left), Aaron D. Bergdahl (middle right), James T. Blodgett (bottom left) and Laurie J. Stepanek (bottom right). To learn more about RMRS publications or search our online titles: www.fs.fed.us/rm/publications www.treesearch.fs.fed.us/ Background This technical report provides a guide to assist arborists, landowners, woody plant pest management specialists, foresters, and plant pathologists in the diagnosis and control of tree diseases encountered in the Great Plains. It contains 56 chapters on tree diseases prepared by 27 authors, and emphasizes disease situations as observed in the 10 states of the Great Plains: Colorado, Kansas, Montana, Nebraska, New Mexico, North Dakota, Oklahoma, South Dakota, Texas, and Wyoming. The need for an updated tree disease guide for the Great Plains has been recog- nized for some time and an account of the history of this publication is provided here. -
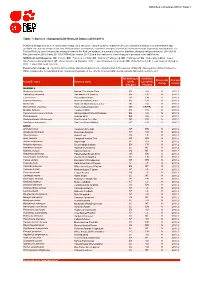
Table 7: Species Changing IUCN Red List Status (2010-2011)
IUCN Red List version 2011.2: Table 7 Table 7: Species changing IUCN Red List Status (2010-2011) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2010 (IUCN Red List version 2010.4) and 2011 (IUCN Red List version 2011.2) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.) IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2010) List (2011) change version Category Category MAMMALS Bradypus torquatus Maned Three-toed Sloth EN VU N 2011.1 Callicebus oenanthe San Martin Titi Monkey EN CR N 2011.1 Equus ferus Przewalski's Horse CR EN G 2011.2 -

European Red List of Non-Marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert
European Red List of Non-marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert European Red List of Non-marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert IUCN Global Species Programme IUCN Regional Office for Europe IUCN Species Survival Commission Published by the European Commission. This publication has been prepared by IUCN (International Union for Conservation of Nature) and the Natural History of Bern, Switzerland. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN, the Natural History Museum of Bern or the European Union concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN, the Natural History Museum of Bern or the European Commission. Citation: Cuttelod, A., Seddon, M. and Neubert, E. 2011. European Red List of Non-marine Molluscs. Luxembourg: Publications Office of the European Union. Design & Layout by: Tasamim Design - www.tasamim.net Printed by: The Colchester Print Group, United Kingdom Picture credits on cover page: The rare “Hélice catalorzu” Tacheocampylaea acropachia acropachia is endemic to the southern half of Corsica and is considered as Endangered. Its populations are very scattered and poor in individuals. This picture was taken in the Forêt de Muracciole in Central Corsica, an occurrence which was known since the end of the 19th century, but was completely destroyed by a heavy man-made forest fire in 2000. -

Discula Lyelliana
The IUCN Red List of Threatened Species™ ISSN 2307-8235 (online) IUCN 2008: T6730A12801850 Discula lyelliana Assessment by: Seddon, M. View on www.iucnredlist.org Citation: Seddon, M. 2013. Discula lyelliana. The IUCN Red List of Threatened Species 2013: e.T6730A12801850. http://dx.doi.org/10.2305/IUCN.UK.2011-1.RLTS.T6730A12801850.en Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale, reposting or other commercial purposes is prohibited without prior written permission from the copyright holder. For further details see Terms of Use. The IUCN Red List of Threatened Species™ is produced and managed by the IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and The IUCN Red List Partnership. The IUCN Red List Partners are: BirdLife International; Botanic Gardens Conservation International; Conservation International; Microsoft; NatureServe; Royal Botanic Gardens, Kew; Sapienza University of Rome; Texas A&M University; Wildscreen; and Zoological Society of London. If you see any errors or have any questions or suggestions on what is shown in this document, please provide us with feedback so that we can correct or extend the information provided. THE IUCN RED LIST OF THREATENED SPECIES™ Taxonomy Kingdom Phylum Class Order Family Animalia Mollusca Gastropoda Stylommatophora Hygromiidae Taxon Name: Discula lyelliana (R.T. Lowe, 1852) Assessment Information Red List Category & Criteria: Critically Endangered (Possibly Extinct) B1ab(iii) ver 3.1 Year Published: 2013 Date Assessed: August 31, 2010 Justification: Seddon (2008) suggested this species should be considered as Critically Endangered B1ab(iii) (version 3.1). -

Porto Santo Candidatura a Reserva Da Biosfera Da UNESCO Março De
Porto Santo Candidatura a Reserva da Biosfera da UNESCO Março de 2019 FICHA TÉCNICA COORDENAÇÃO GERAL Município do Porto Santo Associação Grupo de Folclore do Porto Santo Agência Regional da Energia e Ambiente da Região Autónoma da Madeira Direção Regional para a Administração Pública do Porto Santo Instituto das Florestas e Conservação da Natureza, IP-RAM Secretaria Regional do Ambiente e Recursos Naturais EQUIPA TÉCNICA Duarte Mendonça Filipe Oliveira José Manuel Silva Maria Gorete Freitas Rosa Pires Rubina Brito Rute Areal Susana Fontinha DESIGN E PAGINAÇÃO Neide Paixão Núria Brito FOTOS Dos promotores com exceção das identificadas por: DT – Dinarte Teixeira 4 FV – Filipe Viveiros NP – Neide Paixão NS – Nuno Sá RSM – Rui São Marcos VG – Virgílio Gomes COLABORADORES Ana Gomes Ana Luisa Fernandes António Albuquerque e Silva António Franquinho Aguiar António Iglésias Bruno Cunha Daniel Mata Dília Menezes Dinarte Teixeira Eunice Pinto Francisco Clode Francisco Fernandes Gina Brito Mendes João Batista João Delgado João Rodrigues José Augusto Carvalho José Luís Ferreira Lídia Góes Ferreira 5 Manuel Ara Manuela Sim-Sim Mário Cachão Miguel Ângelo Carvalho Olinda Simone Vasconcelos Raquel Ferreira Ricardo Costa Ricardo Meneses Rui Nunes Sara Andrade Vitor Jorge Vítor Prior AGRADECIMENTOS Direção Regional de Agricultura Direcção Regional da Cultura Direção Regional de Estatística da Madeira Direção Regional de Pescas Direção Regional do Turismo Museu Etnográfico da Madeira Um agradecimento especial aos Porto-santenses que se envolveram neste -

Discula Tectiformis
The IUCN Red List of Threatened Species™ ISSN 2307-8235 (online) IUCN 2008: T6729A12801643 Discula tectiformis Assessment by: Seddon, M.B. View on www.iucnredlist.org Citation: Seddon, M.B. 2013. Discula tectiformis. The IUCN Red List of Threatened Species 2013: e.T6729A12801643. http://dx.doi.org/10.2305/IUCN.UK.2011-1.RLTS.T6729A12801643.en Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale, reposting or other commercial purposes is prohibited without prior written permission from the copyright holder. For further details see Terms of Use. The IUCN Red List of Threatened Species™ is produced and managed by the IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and The IUCN Red List Partnership. The IUCN Red List Partners are: BirdLife International; Botanic Gardens Conservation International; Conservation International; Microsoft; NatureServe; Royal Botanic Gardens, Kew; Sapienza University of Rome; Texas A&M University; Wildscreen; and Zoological Society of London. If you see any errors or have any questions or suggestions on what is shown in this document, please provide us with feedback so that we can correct or extend the information provided. THE IUCN RED LIST OF THREATENED SPECIES™ Taxonomy Kingdom Phylum Class Order Family Animalia Mollusca Gastropoda Stylommatophora Hygromiidae Taxon Name: Discula tectiformis (Sowerby, 1824) Assessment Information Red List Category & Criteria: Endangered B1ab(iii) ver 3.1 Year Published: 2013 Date Assessed: September 30, 2010 Justification: This species has a total extent of occurrence of 42 km2 with the four isolated sub-populations, three on a small hillslope area at the eastern part of the island. -

Impacts of an Exotic Disease and Vegetation Change on Foliar Calcium Cycling in Appalachian Forests
Ecological Applications, 17(3), 2007, pp. 869–881 Ó 2007 by the Ecological Society of America IMPACTS OF AN EXOTIC DISEASE AND VEGETATION CHANGE ON FOLIAR CALCIUM CYCLING IN APPALACHIAN FORESTS 1,4 2 3 MICHAEL A. JENKINS, SHIBU JOSE, AND PETER S. WHITE 1National Park Service, Great Smoky Mountains National Park, Twin Creeks Natural Resources Center, 1314 Cherokee Orchard Road, Gatlinburg, Tennessee 37738 USA 2School of Forest Resources and Conservation, University of Florida, Box 110410, Gainesville, Florida 32611 USA 3Department of Biology–Campus Box 3280, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina 27599-3280 USA Abstract. Because of the high calcium content of its foliage, Cornus florida (flowering dogwood) has been described as a calcium ‘‘pump’’ that draws calcium from deeper mineral soil and enriches surface soil horizons. However, over the last two decades an exotic fungal disease (dogwood anthracnose, Discula destructiva) has decimated populations of this once- common understory species. Its loss, combined with forest stand development, could alter intra-stand calcium cycling. We used data from long-term vegetation monitoring plots to examine the ecological role of C. florida in calcium cycling and to identify changes in annual foliar calcium cycling over a 20-year period between two sampling intervals, 1977–1979 (pre- anthracnose) and 1995–2000 (post-anthracnose). Published equations were used to estimate foliar biomass per species for five forest types: alluvial, typic cove, acid cove, oak–hickory, and oak–pine. Calcium concentrations derived from foliage samples were used to estimate annual foliar calcium production per species for understory woody stems (,20 cm dbh) and total foliar calcium production for overstory stems (20 cm dbh). -

Molecular Phylogeny, Evolution Of
Molecular Phylogeny, Evolution of Shell Shape, and DNA Barcoding in Polygyridae (Gastropoda: Pulmonata), an Endemic North American Clade of Land Snails Author(s): Kathryn E. Perez Nicholas Defreitas , John Slapcinsky , Russell L. Minton , Frank E. Anderson and Timothy A. Pearce Source: American Malacological Bulletin, 32(1):1-31. 2014. Published By: American Malacological Society DOI: http://dx.doi.org/10.4003/006.032.0103 URL: http://www.bioone.org/doi/full/10.4003/006.032.0103 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Amer. Malac. Bull. 32(1): 1–31 (2014) http://www.bioone.org/toc/malb/32/1 Molecular phylogeny, evolution of shell shape, and DNA barcoding in Polygyridae (Gastropoda: Pulmonata), an endemic North American clade of land snails Kathryn E. Perez1, Nicholas Defreitas2, John Slapcinsky3, Russell L. Minton4, Frank E. Anderson2, and Timothy A. -

Lista Dos Moluscos (Mollusca) List of Molluscs
CAPÍTULO 9.1 | CHAPTER 9.1 LISTA DOS MOLUSCOS (MOLLUSCA) LIST OF MOLLUSCS (MOLLUSCA) Cristina Abreu1 & Dinarte Teixeira2 1 Universidade da Madeira, Dep. Biologia/CEM, Edifício da Penteada, 9000 -399 Funchal, Portugal; e -mail: [email protected] 2 Secretaria Regional do Ambiente e dos Recursos Naturais – Direcção Regional do Ambiente – Rua Pestana Júnior nº 6 3º Dto, 9064 -506 Funchal, Madeira, Portugal; e-mail: dinarteteixeira.sra@gov -madeira.pt D MOLLUSCA MA M PS D S Phylum Mollusca Classe Bivalvia Ordem Veneroida Sphaeriidae Pisidium casertanum (Poli, 1791) M Pisidium personatum Malm, 1855 M Classe Gastropoda Ordem Architaenioglossa Cyclophoridae END Craspedopoma lyonnetianum (Lowe, 1852) M END Craspedopoma monizianum Lowe, 1860 M END Craspedopoma mucronatum (Menke, 1830) M END Craspedopoma neritoides Lowe, 1860 M END Craspedopoma trochoideum Lowe, 1860 M Ordem Neotaenioglossa Assimineidae Assiminea eliae Paladilhe, 1875 M Paludinella littorina (Delle Chiaje, 1828) M PS S Hydrobiidae Mercuria balearica (Paladilhe, 1869) M Mercuria similis (Draparnaud, 1805) M Truncatellidae Truncatella subcylindrica (Linnaeus, 1767) M S Ordem Pulmonata Agriolimacidae Deroceras laeve (Muller, 1774) M Deroceras lombricoides (Morelet, 1845) M Deroceras panormitanum (Lessona & Pollonera, 1882) M Deroceras reticulatum (Muller, 1774) M Arionidae Arion hortensis Férussac, 1819 M Arion intermedius Normand, 1852 M Arion lusitanicus Mabille, 1868 M Arion pascalianus Mabille, 1868 M Carychiidae Carychium minimum Muller, 1774 M Carychium tridentatum -

June 2003 Newsletter of the Mycological Society of America
Supplement to Mycologia Vol. 54(3) June 2003 Newsletter of the Mycological Society of America -- In This Issue -- Fungal Bioterrorism Threat Gaining Public Interest, Yet Not Biggest Concern of Fungal Fungal Bioterrorism ................................ 1-2 Specialists, Survey Finds Find of Century: Additional Comments ...... 2 MSA Official Business by Meredith Stone and John Scally From the President .................................. 3 Questions or comments should be sent to John Scally, Senior Account Executive, From the Editor ....................................... 3 G.S. Schwartz & Co. Inc., 470 Park Ave South, 10th Fl. S., New York, NY Mid-Year Executive Council Minutes .. 4-7 10016, 212.725.4500 x 338 or < [email protected] >. Managing Editor’s Mid-Year Report ....... 8 EADING FUNGAL INFECTION EXPERTS to discuss disease challenges Council Email Express ............................. 9 at upcoming mycology medical conference. The threat of fungal Important Announcement .................... 9 Lagents being misused for bioterrorism will gain the most public MSA ABSTRACTS.................. 10-52, 63 attention over the next year, compared with other fungal disease issues, Forms according to one-quarter of fungal (medical mycology) specialists Change of Address ............................... 7 surveyed in an exclusive report. Surprisingly, however, none of those surveyed consider such a bioterrorist threat to be the most significant Endowment & Contributions ............. 64 challenge facing the area of fungal disease. Gift Membership -

Discula Testudinalis, Madeiran Land Snail
The IUCN Red List of Threatened Species™ ISSN 2307-8235 (online) IUCN 2008: T6722A12800171 Discula testudinalis, Madeiran Land Snail Assessment by: Seddon, M.B. View on www.iucnredlist.org Citation: Seddon, M.B. 2013. Discula testudinalis. The IUCN Red List of Threatened Species 2013: e.T6722A12800171. http://dx.doi.org/10.2305/IUCN.UK.2011-1.RLTS.T6722A12800171.en Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale, reposting or other commercial purposes is prohibited without prior written permission from the copyright holder. For further details see Terms of Use. The IUCN Red List of Threatened Species™ is produced and managed by the IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and The IUCN Red List Partnership. The IUCN Red List Partners are: BirdLife International; Botanic Gardens Conservation International; Conservation International; Microsoft; NatureServe; Royal Botanic Gardens, Kew; Sapienza University of Rome; Texas A&M University; Wildscreen; and Zoological Society of London. If you see any errors or have any questions or suggestions on what is shown in this document, please provide us with feedback so that we can correct or extend the information provided. THE IUCN RED LIST OF THREATENED SPECIES™ Taxonomy Kingdom Phylum Class Order Family Animalia Mollusca Gastropoda Stylommatophora Hygromiidae Taxon Name: Discula testudinalis Pfeiffer, 1853 Common Name(s): • English: Madeiran Land Snail Assessment Information Red List Category & Criteria: Critically Endangered B1ab(iii)+2ab(iii) ver 3.1 Year Published: 2013 Date Assessed: September 30, 2010 Justification: This species has been listed since 1980s, and still remains rare, as well as susceptible to predation from non-native species. -

Ornantental Plants Annual Reports and Research Reviews • 1998
Ornantental Plants Annual Reports and Research Reviews • 1998 T·H·E T· H·E OHIO OHIO January 1999 Special Circular 165 m~ Ohio Agricultural Research and Development Center OARilL EXTENSION In Partnership With Ohio State University Extension T · H · E OHIO SfAlE UNIVERSITY OARilL Lowell R. Nault Interim Director Ohio Agricultural Research and Development Center 1680 Madison Avenue Wooster, Ohio 44691-4096 330-263-3700 Ornamental Plants Annual Reports and Research Reviews 1998 Edited By Mary Ann Rose and James A. Chatfield Ohio Agricultural Research and Development Center Ohio State University Extension Department of Horticulture and Crop Science The Ohio State University T · H · E T · H · E OHIO OHIO January 1999 Special Circular 165 srA1EUNIVERSITY SfA1E UNIVERSITY Ohio Agricultural Research and Development Center OARilL EXTENSION In Partnership With Ohio State University Extension 1 Cover Photo: Ken Cochran is shown among the umbrella magnolias (Magnolia tripetla) at the Secrest Arboretum on the Ohio Agricultural Research and Development Center's Wooster campus. Salaries and research support were provided by state and federal funds appropriated to the Ohio Agricultural Research and Development Center and Ohio State University Extension of The Ohio State University's College of Food, Agricultural, and Environmental Sciences. Additional grant support was provided by the organizations and companies listed in the individual research reports. 2 Contents Ohio State University Extension Nursery, Landscape, and Turf Team 7 Directory: 1999 Floriculture Industry Roundtable of Ohio: 1999 12 Ohio State University Extension Buckeye Yard and Garden Line 16 Gary Y. Gao, Pamela J. Bennett, Joseph F. Boggs, James A. Chatfield, David E.