Tracking of Intertissue Migration Reveals the Origins of Tumor-Infiltrating Monocytes
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Development of Plasmacytoid and Conventional Dendritic Cell Subtypes from Single Precursor Cells Derived in Vitro and in Vivo
ARTICLES Development of plasmacytoid and conventional dendritic cell subtypes from single precursor cells derived in vitro and in vivo Shalin H Naik1,2, Priyanka Sathe1,3, Hae-Young Park1,4, Donald Metcalf1, Anna I Proietto1,3, Aleksander Dakic1, Sebastian Carotta1, Meredith O’Keeffe1,4, Melanie Bahlo1, Anthony Papenfuss1, Jong-Young Kwak1,4,LiWu1 & Ken Shortman1 The development of functionally specialized subtypes of dendritic cells (DCs) can be modeled through the culture of bone marrow with the ligand for the cytokine receptor Flt3. Such cultures produce DCs resembling spleen plasmacytoid DCs (pDCs), http://www.nature.com/natureimmunology CD8+ conventional DCs (cDCs) and CD8– cDCs. Here we isolated two sequential DC-committed precursor cells from such cultures: dividing ‘pro-DCs’, which gave rise to transitional ‘pre-DCs’ en route to differentiating into the three distinct DC subtypes (pDCs, CD8+ cDCs and CD8– cDCs). We also isolated an in vivo equivalent of the DC-committed pro-DC precursor cell, which also gave rise to the three DC subtypes. Clonal analysis of the progeny of individual pro-DC precursors demonstrated that some pro-DC precursors gave rise to all three DC subtypes, some produced cDCs but not pDCs, and some were fully committed to a single DC subtype. Thus, commitment to particular DC subtypes begins mainly at this pro-DC stage. Dendritic cells (DCs) are antigen-presenting cells crucial for the innate macrophages12. Further ‘downstream’, ‘immediate’ precursors have and adaptive response to infection as well as for maintaining immune been identified for several DC types, including Ly6Chi monocytes as 3,4,6 13 Nature Publishing Group Group Nature Publishing tolerance to self tissue. -

The Biochemical and Biophysical Mechanisms of Macrophage Migration
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations 2015 The Biochemical and Biophysical Mechanisms of Macrophage Migration Laurel Erin Hind University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Allergy and Immunology Commons, Biomedical Commons, Immunology and Infectious Disease Commons, and the Medical Immunology Commons Recommended Citation Hind, Laurel Erin, "The Biochemical and Biophysical Mechanisms of Macrophage Migration" (2015). Publicly Accessible Penn Dissertations. 1062. https://repository.upenn.edu/edissertations/1062 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/1062 For more information, please contact [email protected]. The Biochemical and Biophysical Mechanisms of Macrophage Migration Abstract The ability of macrophages to migrate is critical for a proper immune response. During an innate immune response, macrophages migrate to sites of infection or inflammation where they clear pathogens through phagocytosis and activate an adaptive immune response by releasing cytokines and acting as antigen- presenting cells. Unfortunately, improper regulation of macrophage migration is associated with a variety of dieases including cancer, atherosclerosis, wound-healing, and rheumatoid arthritis. In this thesis, engineered substrates were used to study the chemical and physical mechanisms of macrophage migration. We first used microcontact printing to generate surfaces -

Single-Cell Mass Cytometry of Differential Immune and Drug
RESEARCH ARTICLE Performance assessmentofmasscytometry. The workflow for mass cytometry is comparable with that of fluorescence flow cytometry (Fig. 1A). Single-Cell Mass Cytometry of Differential Antibodies coupled to distinct, stable, transition element isotopes were used to bind target epitopes on and within cells. Cells, with bound antibody- Immune and Drug Responses Across isotope conjugates, were sprayed as single-cell droplets into an inductively coupled argon plasma a Human Hematopoietic Continuum (created by passing argon gas through an induc- tion coil with a high radio-frequency electric cur- 1 1 2 3 1 1 Sean C. Bendall, * Erin F. Simonds, * Peng Qiu, El-ad D. Amir, Peter O. Krutzik, Rachel Finck, rent) at approximately 5500 K. This vaporizes 1,7 3 1 4,5 6 Robert V. Bruggner, Rachel Melamed, Angelica Trejo, Olga I. Ornatsky, Robert S. Balderas, each cell and induces ionization of its atomic con- 2 1 3 4,5 1 Sylvia K. Plevritis, Karen Sachs, Dana Pe’er, Scott D. Tanner, Garry P. Nolan † stituents. The resulting elemental ions were then sampled by a time-of-flight (TOF) mass spectrom- Flow cytometry is an essential tool for dissecting the functional complexity of hematopoiesis. We used eter and quantified. The signal for each transi- single-cell “mass cytometry” to examine healthy human bone marrow, measuring 34 parameters tion element isotope reporter was integrated as simultaneously in single cells (binding of 31 antibodies, viability, DNA content, and relative cell size). The each cell’s constituent ions reached the detector. signaling behavior of cell subsets spanning a defined hematopoietic hierarchy was monitored with 18 Currently, TOF sampling resolution enables the simultaneous markers of functional signaling states perturbed by a set of ex vivo stimuli and inhibitors. -

Monocyte and Macrophage Heterogeneity
REVIEWS MONOCYTE AND MACROPHAGE HETEROGENEITY Siamon Gordon and Philip R. Taylor Abstract | Heterogeneity of the macrophage lineage has long been recognized and, in part, is a result of the specialization of tissue macrophages in particular microenvironments. Circulating monocytes give rise to mature macrophages and are also heterogeneous themselves, although the physiological relevance of this is not completely understood. However, as we discuss here, recent studies have shown that monocyte heterogeneity is conserved in humans and mice, allowing dissection of its functional relevance: the different monocyte subsets seem to reflect developmental stages with distinct physiological roles, such as recruitment to inflammatory lesions or entry to normal tissues. These advances in our understanding have implications for the development of therapeutic strategies that are targeted to modify particular subpopulations of monocytes. OSTEOCLAST Circulating monocytes give rise to a variety of tissue- elicit increased recruitment of monocytes to peripheral A multinucleate cell that resident macrophages throughout the body, as well as sites4, where differentiation into macrophages and DCs resorbs bone. to specialized cells such as dendritic cells (DCs) and occurs, contributing to host defence, and tissue remod- OSTEOCLASTS. Monocytes are known to originate in the elling and repair. Studies of the mononuclear-phagocyte bone marrow from a common myeloid progenitor that system, using monoclonal antibodies specific for vari- is shared with neutrophils, and they are then released ous cell-surface receptors and differentiation antigens, into the peripheral blood, where they circulate for have shown that there is substantial heterogeneity of several days before entering tissues and replenishing phenotype, which most probably reflects the special- the tissue macrophage populations1. -
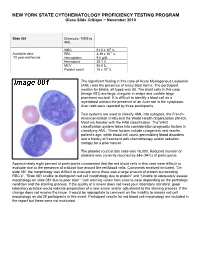
10 11 Cyto Slides 81-85
NEW YORK STATE CYTOHEMATOLOGY PROFICIENCY TESTING PROGRAM Glass Slide Critique ~ November 2010 Slide 081 Diagnosis: MDS to AML 9 WBC 51.0 x 10 /L 12 Available data: RBC 3.39 x 10 /L 72 year-old female Hemoglobin 9.6 g/dL Hematocrit 29.1 % MCV 86.0 fL Platelet count 16 x 109 /L The significant finding in this case of Acute Myelogenous Leukemia (AML) was the presence of many blast forms. The participant median for blasts, all types was 88. The blast cells in this case (Image 081) are large, irregular in shape and contain large prominent nucleoli. It is difficult to identify a blast cell as a myeloblast without the presence of an Auer rod in the cytoplasm. Auer rods were reported by three participants. Two systems are used to classify AML into subtypes, the French- American-British (FAB) and the World Health Organization (WHO). Most are familiar with the FAB classification. The WHO classification system takes into consideration prognostic factors in classifying AML. These factors include cytogenetic test results, patient’s age, white blood cell count, pre-existing blood disorders and a history of treatment with chemotherapy and/or radiation therapy for a prior cancer. The platelet count in this case was 16,000. Reduced number of platelets was correctly reported by 346 (94%) of participants. Approximately eight percent of participants commented that the red blood cells in this case were difficult to evaluate due to the presence of a bluish hue around the red blood cells. Comments received included, “On slide 081 the morphology was difficult to evaluate since there was a large amount of protein surrounding RBC’s”, “Slide 081 unable to distinguish red cell morphology due to protein” and “Unable to adequately assess morphology on slide 081 due to poor stain”. -

10 Maturation and Development of Leucocytes
MODULE Maturation and Development of Leucocytes Hematology and Blood Bank Technique 10 Notes MATURATION AND DEVELOPMENT OF LEUCOCYTES 10.1 INTRODUCTION The leucocytes develop from the multipotent hematopoietic stem cell which then gives rise to a stem cell committed to formation of leucocytes. Both these cells cannot be identified morphologically by routine methods. The various types of leucocytes are granulocytes (neutrophils, eosinophils and basophils), monocytes and lymphocytes. The three cell types develop separately and accordingly these processes will be discussed separately. OBJECTIVES After reading this lesson, you will be able to: z explain the various stages in the development of leucocytes. z describe the different types of leucocytes seen normally in PBF. 10.2 MYELOPOIESIS This is the process of formation of myeloid cells. It is restricted to the bone marrow after birth. The committed progenitor cell for granulocytes and monocytes is the GM-CFU which proliferates and differentiates to form myeloblast and monoblast. The myeloblast is the earliest morphologically identifiable cell. It is 10-18µm in size. The cytoplasm is scant and basophilic, usually agranular and may contain a few azurophilic cytoplasmic granules in the blast transiting to the next stage 80 HEMATOLOGY AND BLOOD BANK TECHNIQUE Maturation and Development of Leucocytes MODULE of promyelocyte. It has a large round to oval nucleus with a smooth nuclear Hematology and Blood membrane. The chromatin is fine, lacy and is evenly distributed throughout the Bank Technique nucleus. Two-five nucleoli can be identified in the nucleus. The next stage of maturation is the Promyelocyte. It is larger than a myeloblast, 12-20 µm with more abundant cytoplasm which has abundant primary azurophilic granules . -

Acute Monoblastic Leukemia with T(10;11)(P12;Q23) Presenting with Pulmonary Involvement: a Case Report and Literature Review
Case Report Acute Monoblastic Leukemia with t(10;11)(p12;q23) Presenting with Pulmonary Involvement: A Case Report and Literature Review Kannadit Prayongratana MD*, Manaphol Kulpraneet MD*, Prapaporn Panichchob BSc**, Woraphot Tantisiriwat MD*** * Department of Medicine, Faculty of Medicine, Srinakharinwirot University, Nakhon-nayok ** Department of Pathology, Faculty of Medicine, Srinakharinwirot University, Nakhon-nayok *** Department of Preventive and Social Medicine. Faculty of Medicine, Srinakharinwirot University, Nakhon-nayok A forty-three-year-old Thai man presented with acute fever and dyspnea for one week with bilateral patchy infiltration, pancytopenia with monoblast. Bone marrow study was consistent with acute monoblastic leukemia. Lung lesions rapidly progressed to acute respiratory failure, which required intubation. Broncho- scopy with bronchoalveolar lavage revealed monotonous monoblast infiltration. Induction chemotherapy with 7 + 3 regimen was administered to halt the progression of leukemic pulmonary infiltration. Although there was clinical improvement, the chest radiograph developed crescent formation in the right upper lung field. Invasive pulmonary aspergillosis was suspected and successfully treated with antifungal agent. After peripheral blood recovery, bone marrow evaluation was performed and complete remission was established. HLA matching was sent to prepare for hematopoietic stem cell transplantation (HSCT). The literature review showed that the appropriate treatment for the patients with t(10;11)(p12;q23) was HSCT, but there was no data concerning correlation of t(10;11)(p12;q23) and pulmonary infiltration. This may be due to the low incidence of leukemic infiltration of acute leukemia patients, which is 0.48% and 3.06% in acute myeloid leukemia and acute monoblastic leukemia, respectively. Keywords: Acute monoblastic leukemia, Leukemic pulmonary infiltration, t(10;11)(p12;q23) J Med Assoc Thai 2008; 91 (4): 559-63 Full text. -

Sysmex SEED H Monocyte Counting
SYSMEX EDUCATIONAL ENHANCEMENT AND DEVELOPMENT | NOVEMBER 2016 SEED HAEMATOLOGY The blast cell – a diagnostic heavyweight Causes and cytological manifestations Blast cells are described as precursor cells with the ability to preserve themselves by dividing and to further differentiate. Under pathological conditions, blast cells can be mobilised from the bone marrow into the peripheral blood circulation. In adults, this represents an alarming finding that can indicate both reactive and malignant diseases such as leukaemia. Therefore the detection of blast cells in the peripheral blood is considered extremely important, and great responsibility is placed on the investigating laboratory. As well as informa- Fig. 1 Asymmetrical replication (example erythropoiesis) tion on the physiology, this article describes the possible causes of the release of blast cells into the blood, the char- A mature cell is developed after several differentiation acteristics by which they can be identified and how further stages, involving gradual condensation of the nuclear chro- diagnosis is carried out. matin. While blast cells have a homogeneous chromatin, the nucleus shows chromatin clumping in the mature cells. The Development, maturation and regulation nucleus-plasma relation also drops (see Fig. 2) Haematopoietic precursor cells develop from the pluripotent embryonic stem cells as a result of numerous development stages. In the bone marrow, these cells are referred to as blast Maturation cells (‘blastós’ is the Greek word for germ, bud, sprout or shoot). For their further development, they are committed to one specific line (erythropoiesis, granulopoiesis, monopoiesis, Blast cell Mature cell Nuclear chromatin fine, homogeneous Nuclear chromatin clumped thrombopoiesis and lymphopoiesis). Asymmetrical replication, Nucleus-plasma relation 70 – 95 % Nucleus-plasma relation 30 – 50 % as shown in Fig. -

Hemopoietic Progenitors + Differentiation of Human CD34
The Journal of Immunology The Vitamin D3/Hox-A10 Pathway Supports MafB Function during the Monocyte Differentiation of Human CD34؉ Hemopoietic Progenitors1 Claudia Gemelli, Claudia Orlandi, Tommaso Zanocco Marani, Andrea Martello, Tatiana Vignudelli, Francesco Ferrari, Monica Montanari, Sandra Parenti, Anna Testa, Alexis Grande,2,3 and Sergio Ferrari2 Although a considerable number of reports indicate an involvement of the Hox-A10 gene in the molecular control of hemopoiesis, the conclusions of such studies are quite controversial given that they support, in some cases, a role in the stimulation of stem cell self-renewal and myeloid progenitor expansion, whereas in others they implicate this transcription factor in the induction of monocyte-macrophage differentiation. To clarify this issue, we analyzed the biological effects and the transcriptome changes determined in human primary CD34؉ hemopoietic progenitors by retroviral transduction of a full-length Hox-A10 cDNA. The results obtained clearly indicated that this homeogene is an inducer of monocyte differentiation, at least partly acting through the up-regulation of the MafB gene, recently identified as the master regulator of such a maturation pathway. By using a combined approach based on computational analysis, EMSA experiments, and luciferase assays, we were able to demonstrate the presence of a Hox-A10-binding site in the promoter region of the MafB gene, which suggested the likely molecular mechanism underlying the observed effect. Stimulation of the same cells with the vitamin D3 monocyte differentiation inducer resulted in a clear increase of Hox-A10 and MafB transcripts, indicating the existence of a precise transactivation cascade involving vitamin D3 receptor, Hox-A10, and MafB transcription factors. -

Regulation of the Proliferation of the Established Human Monoblast Cell Line, U937, at the Single Cell Level1
[CANCER RESEARCH 46, 3309-3312, July 1986] Regulation of the Proliferation of the Established Human Monoblast Cell Line, U937, at the Single Cell Level1 Ellin Berman, Berish Y. Rubin, and Hal E. Broxmeyer2 Department of Developmental Hematopoiesis, Memorial Sloan Kettering Cancer Center [E. B.J; Department of Lymphokine Biology, The Lindsley F. Kimball Research Institute of the New York Blood Center, New York, New York 10021 [B. Y. R.J; and Departments of Medicine, Hematology /Oncology, Microbiology and Immunology, Indiana University School of Medicine, Walther Medical Research Institute and the Elks Cancer Research Center, Indianapolis, Indiana 46223 ¡H.E. B.] ABSTRACT pliages and T-lymphocytes (4, 12). Colony formation by MHC class II antigen-positive U937 CFC can be suppressed by puri U937 cells, an established monoblast or early monocyte cell line, were fied iron-saturated LF and TF and this is associated with assessed as a model in vitro for the regulation of cell growth at the single induction of MHC class II antigens on U937 CFC by HuIFN- cell level. Colony formation by 500 U937 cells, preinduced to a state of •y(13,14). Dialyzed U937 CM does not influence the cloning responsiveness to lactoferrin (LF) by incubation with human y Interferon was suppressed by LF. LF-suppressed colony formation was restored by efficiency of U937 cells but does restore colony formation partially purified growth activity derived from U937 cells. The release of suppressed by LF or TF without inactivating the LF or TF growth factor(s) into conditioned medium required concentrations of >51III (15). These results were similar to studies reported previously U937 cells/ml and this release was dependent on the length of time that with a murine myelomonocytic leukemia cell line, WEHI-3, the cells conditioned the culture medium. -

A Peculiar Case of Acute Myeloid Leukemia Mimicking Plasmacytoid Dendritic Precursor Cell Leukemia
J Clin Exp Hematopathol Vol. 48, No. 2, Nov. 2008 Case Study A Peculiar Case of Acute Myeloid Leukemia Mimicking Plasmacytoid Dendritic Precursor Cell Leukemia Fuminori Sano,1) Taizo Tasaka,2) Hirotake Nishimura,3) Takashi Akiyama,3) Yasutaka Kubo,1) Yoshiko Matsuhashi,1) Hideho Wada,1) Takashi Sugihara,1) Mitsunori Yamakawa,4) and Yoshito Sadahira3) Differential diagnosis between plasmacytoid dendritic precursor cell leukemia (pDC leukemia) and acute myeloid leukemia (AML) with monocytic differentiation is difficult due to shared clinicopathological features ; however, such diagnosis is critical because the two leukemias are treated differently. Here we report a peculiar case of AML mimicking pDC leukemia. A 22- year-old man presented with leukocytopenia and bone marrow involvement of atypical plasmacytoid cells with a prominent nucleolus. In spite of positive cytochemical staining for NaF-sensitive naphthyl butyrate esterase, this case was diagnosed as pDC leukemia because the abnormal cells were positive for CD4, CD56, and CD123, and negative for myeloperoxidase and lysozyme. The patient achieved complete remission after 4 courses of combination chemotherapy, but relapsed four months later with leukemic manifestation and skin involvement. The morphology of the leukemia cells became myelomonoblastic, and some were immunohistochemically positive for lysozyme, suggesting AML. Although the patient received allogenic stem cell transplantation twice, he died of progressive disease. This case demonstrates the importance of cytochemical staining for naphthyl butyrate esterase in differential diagnosis between AML and pDC leukemia coexpressing CD4, CD56, and CD123. 〔J Clin Exp Hematopathol 48(2) : 65- 69, 2008〕 Keywords: dendritic precursor cell leukemia, acute myeloid leukemia, naphthyl butyrate esterase staining and negative for the myeloid markers CD14 and CD16, the T- INTRODUCTION cell marker CD3, and the B-cell markers CD19 and CD20. -

Monocytes, Macrophages, and Their Diseases in Man
oo22-202X/ 78/ 7101-0056$02.oo/ 0 THE Jo fiNA L O F INVESTIGATIVE DERMATOLOGY, 71:56-58,1978 Vol. 71, NO.1 Copyright © 1978 by The Williams & Wilkins Co. Printed in U.S.A. I Monocytes, Macrophages, and Their Diseases in Man MARTiN J. CLINE, M.D. Division of Hematology-Oncology, Department of Medicine, University of California School of Medicine, Los Angeles, California, U.S.A. Mononuclear phagocytes are a widely distributed sys pool in a ratio of 1:3, (iii) monocytes leave the blood exponen tem of cells involved in host defense functions, immu tially with a half life of 8.4 hr, (iv), the half life may be prolonged nologic reactions, disposal of unwanted organic and in in patients with monocytosis and shortened in patients with \ organic materials, and synthesis of biologically active acute infection or splenomegaly, and (v) the normal monocyte compounds such as complement components and pros turnover rate averages 7 X lOG cells pel' hour pel' kilogram of taglandins. The system includes early precursor cells in body weight. the bone marrow, blood monocytes, alveolar and peri Available data suggest that there is no significant reentry of toneal macrophages, sinusoidal lining cells in the spleen, monocytes into the bloodstream once they have left the circu and Kupffer cells in the liver. We have recently come to lation. They enter the tissues and mature to macrophages. The understand the biology of these interesting cells and the major source of mononuclear phagocytes accumulating at in diseases they produce. flammatory foci is the monocyte precursors of the bone marrow [6].