The Biochemical and Biophysical Mechanisms of Macrophage Migration
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Regulation of Macrophage Development and Function in Peripheral Tissues
REVIEWS Regulation of macrophage development and function in peripheral tissues Yonit Lavin, Arthur Mortha, Adeeb Rahman and Miriam Merad Abstract | Macrophages are immune cells of haematopoietic origin that provide crucial innate immune defence and have tissue-specific functions in the regulation and maintenance of organ homeostasis. Recent studies of macrophage ontogeny, as well as transcriptional and epigenetic identity, have started to reveal the decisive role of the tissue stroma in the regulation of macrophage function. These findings suggest that most macrophages seed the tissues during embryonic development and functionally specialize in response to cytokines and metabolites that are released by the stroma and drive the expression of unique transcription factors. In this Review, we discuss how recent insights into macrophage ontogeny and macrophage–stroma interactions contribute to our understanding of the crosstalk that shapes macrophage function and the maintenance of organ integrity. Mononuclear phagocyte Macrophages are key components of the innate immune characterized the transcriptional and epigenetic pro- system system that reside in tissues, where they function as grammes of tissue-resident macrophages and revealed (MPS). A group of bone immune sentinels. They are uniquely equipped to sense the extent of diversity in these populations1,8. In addi- marrow-derived cells and respond to tissue invasion by infectious microorgan- tion to differences in ontogeny, locally derived tissue (monocytes, macrophages and isms and tissue -

Development of Plasmacytoid and Conventional Dendritic Cell Subtypes from Single Precursor Cells Derived in Vitro and in Vivo
ARTICLES Development of plasmacytoid and conventional dendritic cell subtypes from single precursor cells derived in vitro and in vivo Shalin H Naik1,2, Priyanka Sathe1,3, Hae-Young Park1,4, Donald Metcalf1, Anna I Proietto1,3, Aleksander Dakic1, Sebastian Carotta1, Meredith O’Keeffe1,4, Melanie Bahlo1, Anthony Papenfuss1, Jong-Young Kwak1,4,LiWu1 & Ken Shortman1 The development of functionally specialized subtypes of dendritic cells (DCs) can be modeled through the culture of bone marrow with the ligand for the cytokine receptor Flt3. Such cultures produce DCs resembling spleen plasmacytoid DCs (pDCs), http://www.nature.com/natureimmunology CD8+ conventional DCs (cDCs) and CD8– cDCs. Here we isolated two sequential DC-committed precursor cells from such cultures: dividing ‘pro-DCs’, which gave rise to transitional ‘pre-DCs’ en route to differentiating into the three distinct DC subtypes (pDCs, CD8+ cDCs and CD8– cDCs). We also isolated an in vivo equivalent of the DC-committed pro-DC precursor cell, which also gave rise to the three DC subtypes. Clonal analysis of the progeny of individual pro-DC precursors demonstrated that some pro-DC precursors gave rise to all three DC subtypes, some produced cDCs but not pDCs, and some were fully committed to a single DC subtype. Thus, commitment to particular DC subtypes begins mainly at this pro-DC stage. Dendritic cells (DCs) are antigen-presenting cells crucial for the innate macrophages12. Further ‘downstream’, ‘immediate’ precursors have and adaptive response to infection as well as for maintaining immune been identified for several DC types, including Ly6Chi monocytes as 3,4,6 13 Nature Publishing Group Group Nature Publishing tolerance to self tissue. -

"Macrophage Function Disorders". In: Encyclopedia of Life Sciences (ELS)
Macrophage Function Advanced article Disorders Article Contents . Introduction Keith M Lee, McMaster Immunology Research Centre & M. G. DeGroote Institute for . Macrophage Functions . Macrophage Phenotypic Diversity Infectious Disease Research, McMaster University, Hamilton, Ontario, Canada . Role in Disease Charles Yin, McMaster Immunology Research Centre & M. G. DeGroote Institute for Infectious . Primary Immunodeficiencies in Macrophage Function Disease Research, McMaster University, Hamilton, Ontario, Canada . Concluding Remarks Chris P Verschoor, McMaster Immunology Research Centre & M. G. DeGroote Institute for Online posting date: 20th September 2013 Infectious Disease Research, McMaster University, Hamilton, Ontario, Canada Dawn ME Bowdish, McMaster Immunology Research Centre & M. G. DeGroote Institute for Infectious Disease Research, McMaster University, Hamilton, Ontario, Canada Based in part on the previous version of this eLS article ‘Macrophage Function Disorders’ (2009) by Dawn ME Bowdish and Siamon Gordon. Macrophages are sentinel cells of the innate immune tissue microenvironment and can change considerably response. Macrophages recognise pathogen-associated with exposure to infectious and antigenic agents. They are molecular patterns (e.g. microbial products) and endo- relatively long-lived, biosynthetically active cells and genous ligands (e.g. apoptotic cells) through a broad and express diverse surface receptors and secretory products. They adapt readily to changes in their milieu and help to adaptable range of pattern-recognition receptors. The maintain homoeostasis locally and systemically. If unable consequence of this recognition is generally effective to deal adequately with an infectious or injurious stimulus, clearance via phagocytosis; however, when this is not macrophages initiate a chronic inflammatory process, effective, macrophages may become inappropriately which can contribute to persistent tissue damage; they can activated and initiate an inappropriate inflammatory also mediate acute, sometimes massive, responses from response. -

Understanding the Immune System: How It Works
Understanding the Immune System How It Works U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES NATIONAL INSTITUTES OF HEALTH National Institute of Allergy and Infectious Diseases National Cancer Institute Understanding the Immune System How It Works U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES NATIONAL INSTITUTES OF HEALTH National Institute of Allergy and Infectious Diseases National Cancer Institute NIH Publication No. 03-5423 September 2003 www.niaid.nih.gov www.nci.nih.gov Contents 1 Introduction 2 Self and Nonself 3 The Structure of the Immune System 7 Immune Cells and Their Products 19 Mounting an Immune Response 24 Immunity: Natural and Acquired 28 Disorders of the Immune System 34 Immunology and Transplants 36 Immunity and Cancer 39 The Immune System and the Nervous System 40 Frontiers in Immunology 45 Summary 47 Glossary Introduction he immune system is a network of Tcells, tissues*, and organs that work together to defend the body against attacks by “foreign” invaders. These are primarily microbes (germs)—tiny, infection-causing Bacteria: organisms such as bacteria, viruses, streptococci parasites, and fungi. Because the human body provides an ideal environment for many microbes, they try to break in. It is the immune system’s job to keep them out or, failing that, to seek out and destroy them. Virus: When the immune system hits the wrong herpes virus target or is crippled, however, it can unleash a torrent of diseases, including allergy, arthritis, or AIDS. The immune system is amazingly complex. It can recognize and remember millions of Parasite: different enemies, and it can produce schistosome secretions and cells to match up with and wipe out each one of them. -

Infections Macrophage Polarization in Bacterial
Macrophage Polarization in Bacterial Infections Marie Benoit, Benoît Desnues and Jean-Louis Mege This information is current as J Immunol 2008; 181:3733-3739; ; of September 24, 2021. doi: 10.4049/jimmunol.181.6.3733 http://www.jimmunol.org/content/181/6/3733 Downloaded from References This article cites 68 articles, 21 of which you can access for free at: http://www.jimmunol.org/content/181/6/3733.full#ref-list-1 Why The JI? Submit online. http://www.jimmunol.org/ • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists • Fast Publication! 4 weeks from acceptance to publication *average by guest on September 24, 2021 Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2008 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. Macrophage Polarization in Bacterial Infections Marie Benoit, Benoît Desnues, and Jean-Louis Mege1 Converging studies have shown that M1 and M2 mac- tokines and microbial products (2). More recently, M2 macro- rophages are functionally polarized in response to mi- phages have been characterized by functional expression of al- croorganisms and host mediators. Gene expression ternative activation markers. -

Granulocytes, Macrophages, and Dendritic Cells Arise from A
Proc. Natl. Acad. Sci. USA Vol. 90, pp. 3038-3042, April 1993 Immunology Granulocytes, macrophages, and dendritic cells arise from a common major histocompatibility complex class II-negative progenitor in mouse bone marrow KAYO INABA*t, MUNEO INABA*, MASASHI DEGUCHI*, KATSUHIKO HAGI*, RYoJi YASUMIZUf, SUSUMU IKEHARAt, SHIGERU MURAMATSU*, AND RALPH M. STEINMAN§ *Department of Zoology, Faculty of Science, Kyoto University, Sakyo, Kyoto 606, Japan; tFirst Department of Pathology, Kansai Medical University, Moriguchi, Osaka 570, Japan; and §Laboratory of Cellular Physiology and Immunology, The Rockefeller University, New York, NY 10021 Communicated by Zanvil A. Cohn, December 21, 1992 ABSTRACT The developmental origin of dendritic cells, a lineage-restricted macrophage and granulocyte colony- specialized system ofmajor histocompatibility complex (MHC) stimulating factors (M-CSF and G-CSF, respectively) (8, 10, class 11-rich antigen-presenting cells for T-celi immunity and 12). tolerance, is not well characterized. Granulocyte-macrophage Since GM-CSF can induce the formation of mixed popu- colony-stimulating factor (GM-CSF) is known to stimulate lations ofgranulocytes and macrophages in semi-solid colony dendritic cells, including growth and development from MHC systems (15), we asked whether dendritic cells could also class 11-negative precursors in suspension cultures of mouse arise from a colony-forming precursor that is common to bone marrow. Here we studied colony formation in semi-solid phagocytes. Cells with some of the features of dendritic cells methylcellulose cultures, a classical bioassay system in which have been detected in human cell colonies that were induced GM-CSF induces the formation of mixed granulocyte- with lectin-conditioned medium (16) and more recently with macrophage colonies. -

Ornithine Decarboxylase Regulates M1 Macrophage Activation And
Ornithine decarboxylase regulates M1 macrophage PNAS PLUS activation and mucosal inflammation via histone modifications Dana M. Hardbowera,b, Mohammad Asimb, Paula B. Luisc, Kshipra Singhb, Daniel P. Barryb, Chunying Yangd,e, Meredith A. Steevese, John L. Clevelandd,e, Claus Schneiderc, M. Blanca Piazuelob, Alain P. Gobertb, and Keith T. Wilsona,b,f,g,h,1 aDepartment of Pathology, Microbiology and Immunology, Vanderbilt University Medical Center, Nashville, TN 37232; bDivision of Gastroenterology, Hepatology and Nutrition, Department of Medicine, Vanderbilt University Medical Center, Nashville, TN 37232; cDepartment of Pharmacology, Vanderbilt University School of Medicine, Nashville, TN 37232; dDepartment of Tumor Biology, Moffitt Cancer Center and Research Institute, Tampa, FL 33612; eDepartment of Cancer Biology, The Scripps Research Institute, Jupiter, FL 33458; fDepartment of Cancer Biology, Vanderbilt University School of Medicine, Nashville, TN 37232; gCenter for Mucosal Inflammation and Cancer, Vanderbilt University Medical Center, Nashville, TN 37232; and hVeterans Affairs Tennessee Valley Healthcare System, Nashville, TN 37232 Edited by Carl F. Nathan, Weill Medical College of Cornell University, New York, NY, and approved December 20, 2016 (received for review September 6, 2016) Macrophage activation is a critical step in host responses during pathogens is macrophage activation. Macrophages have the ca- bacterial infections. Ornithine decarboxylase (ODC), the rate-limiting pacity to alter cytokine/chemokine production and various other enzyme in polyamine metabolism, has been well studied in epithelial functions along the spectrum of M1 or M2 activation based on the cells and is known to have essential roles in many different cellular stimulus detected (12–14). M1 macrophages represent a highly functions. However, its role in regulating macrophage function during proinflammatory and antimicrobial subset of macrophages (12–14). -

Cd47-Sirpα Interaction and IL-10 Constrain Inflammation-Induced Macrophage Phagocytosis of Healthy Self-Cells
Cd47-Sirpα interaction and IL-10 constrain inflammation-induced macrophage phagocytosis of healthy self-cells Zhen Biana,b, Lei Shia, Ya-Lan Guoa, Zhiyuan Lva, Cong Tanga, Shuo Niua, Alexandra Tremblaya, Mahathi Venkataramania, Courtney Culpeppera, Limin Lib, Zhen Zhoub, Ahmed Mansoura, Yongliang Zhangc, Andrew Gewirtzd, Koby Kiddera,e, Ke Zenb, and Yuan Liua,d,1 aProgram of Immunology and Cell Biology, Department of Biology, Center for Diagnostics & Therapeutics, Georgia State University, Atlanta, GA 30302; bState Key Laboratory of Pharmaceutical Biotechnology, Nanjing Advanced Institute for Life Sciences, Nanjing University, Nanjing, Jiangsu 210093, China; cDepartment of Microbiology and Immunology, Yong Loo Lin School of Medicine, Life Science Institute (LSI) Immunology Programme, National University of Singapore, Singapore 117456; dCenter for Inflammation, Immunity and Infection, Georgia State University, Atlanta, GA 30303; and eDepartment of Cell Biology, Rutgers University, New Brunswick, NJ 08901 Edited by Jason G. Cyster, University of California, San Francisco, CA, and approved July 11, 2016 (received for review October 28, 2015) − − Rapid clearance of adoptively transferred Cd47-null (Cd47 / ) cells in The CD47-SIRPα mechanism was first reported by Oldenborg congeneic WT mice suggests a critical self-recognition mechanism, et al. (1), who had demonstrated in red blood cell (RBC) in which CD47 is the ubiquitous marker of self, and its interaction transfusion experiments that WT mice rapidly eliminate syngeneic − with macrophage signal regulatory protein α (SIRPα) triggers inhib- Cd47-null (Cd47 ) RBCs through erythrophagocytosis in the itory signaling through SIRPα cytoplasmic immunoreceptor tyrosine- spleen and that the lack of tyrosine phosphorylation in SIRPα based inhibition motifs and tyrosine phosphatase SHP-1/2. -

Single-Cell Mass Cytometry of Differential Immune and Drug
RESEARCH ARTICLE Performance assessmentofmasscytometry. The workflow for mass cytometry is comparable with that of fluorescence flow cytometry (Fig. 1A). Single-Cell Mass Cytometry of Differential Antibodies coupled to distinct, stable, transition element isotopes were used to bind target epitopes on and within cells. Cells, with bound antibody- Immune and Drug Responses Across isotope conjugates, were sprayed as single-cell droplets into an inductively coupled argon plasma a Human Hematopoietic Continuum (created by passing argon gas through an induc- tion coil with a high radio-frequency electric cur- 1 1 2 3 1 1 Sean C. Bendall, * Erin F. Simonds, * Peng Qiu, El-ad D. Amir, Peter O. Krutzik, Rachel Finck, rent) at approximately 5500 K. This vaporizes 1,7 3 1 4,5 6 Robert V. Bruggner, Rachel Melamed, Angelica Trejo, Olga I. Ornatsky, Robert S. Balderas, each cell and induces ionization of its atomic con- 2 1 3 4,5 1 Sylvia K. Plevritis, Karen Sachs, Dana Pe’er, Scott D. Tanner, Garry P. Nolan † stituents. The resulting elemental ions were then sampled by a time-of-flight (TOF) mass spectrom- Flow cytometry is an essential tool for dissecting the functional complexity of hematopoiesis. We used eter and quantified. The signal for each transi- single-cell “mass cytometry” to examine healthy human bone marrow, measuring 34 parameters tion element isotope reporter was integrated as simultaneously in single cells (binding of 31 antibodies, viability, DNA content, and relative cell size). The each cell’s constituent ions reached the detector. signaling behavior of cell subsets spanning a defined hematopoietic hierarchy was monitored with 18 Currently, TOF sampling resolution enables the simultaneous markers of functional signaling states perturbed by a set of ex vivo stimuli and inhibitors. -

Monocyte and Macrophage Heterogeneity
REVIEWS MONOCYTE AND MACROPHAGE HETEROGENEITY Siamon Gordon and Philip R. Taylor Abstract | Heterogeneity of the macrophage lineage has long been recognized and, in part, is a result of the specialization of tissue macrophages in particular microenvironments. Circulating monocytes give rise to mature macrophages and are also heterogeneous themselves, although the physiological relevance of this is not completely understood. However, as we discuss here, recent studies have shown that monocyte heterogeneity is conserved in humans and mice, allowing dissection of its functional relevance: the different monocyte subsets seem to reflect developmental stages with distinct physiological roles, such as recruitment to inflammatory lesions or entry to normal tissues. These advances in our understanding have implications for the development of therapeutic strategies that are targeted to modify particular subpopulations of monocytes. OSTEOCLAST Circulating monocytes give rise to a variety of tissue- elicit increased recruitment of monocytes to peripheral A multinucleate cell that resident macrophages throughout the body, as well as sites4, where differentiation into macrophages and DCs resorbs bone. to specialized cells such as dendritic cells (DCs) and occurs, contributing to host defence, and tissue remod- OSTEOCLASTS. Monocytes are known to originate in the elling and repair. Studies of the mononuclear-phagocyte bone marrow from a common myeloid progenitor that system, using monoclonal antibodies specific for vari- is shared with neutrophils, and they are then released ous cell-surface receptors and differentiation antigens, into the peripheral blood, where they circulate for have shown that there is substantial heterogeneity of several days before entering tissues and replenishing phenotype, which most probably reflects the special- the tissue macrophage populations1. -
10 11 Cyto Slides 81-85
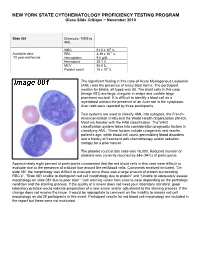
NEW YORK STATE CYTOHEMATOLOGY PROFICIENCY TESTING PROGRAM Glass Slide Critique ~ November 2010 Slide 081 Diagnosis: MDS to AML 9 WBC 51.0 x 10 /L 12 Available data: RBC 3.39 x 10 /L 72 year-old female Hemoglobin 9.6 g/dL Hematocrit 29.1 % MCV 86.0 fL Platelet count 16 x 109 /L The significant finding in this case of Acute Myelogenous Leukemia (AML) was the presence of many blast forms. The participant median for blasts, all types was 88. The blast cells in this case (Image 081) are large, irregular in shape and contain large prominent nucleoli. It is difficult to identify a blast cell as a myeloblast without the presence of an Auer rod in the cytoplasm. Auer rods were reported by three participants. Two systems are used to classify AML into subtypes, the French- American-British (FAB) and the World Health Organization (WHO). Most are familiar with the FAB classification. The WHO classification system takes into consideration prognostic factors in classifying AML. These factors include cytogenetic test results, patient’s age, white blood cell count, pre-existing blood disorders and a history of treatment with chemotherapy and/or radiation therapy for a prior cancer. The platelet count in this case was 16,000. Reduced number of platelets was correctly reported by 346 (94%) of participants. Approximately eight percent of participants commented that the red blood cells in this case were difficult to evaluate due to the presence of a bluish hue around the red blood cells. Comments received included, “On slide 081 the morphology was difficult to evaluate since there was a large amount of protein surrounding RBC’s”, “Slide 081 unable to distinguish red cell morphology due to protein” and “Unable to adequately assess morphology on slide 081 due to poor stain”. -

10 Maturation and Development of Leucocytes
MODULE Maturation and Development of Leucocytes Hematology and Blood Bank Technique 10 Notes MATURATION AND DEVELOPMENT OF LEUCOCYTES 10.1 INTRODUCTION The leucocytes develop from the multipotent hematopoietic stem cell which then gives rise to a stem cell committed to formation of leucocytes. Both these cells cannot be identified morphologically by routine methods. The various types of leucocytes are granulocytes (neutrophils, eosinophils and basophils), monocytes and lymphocytes. The three cell types develop separately and accordingly these processes will be discussed separately. OBJECTIVES After reading this lesson, you will be able to: z explain the various stages in the development of leucocytes. z describe the different types of leucocytes seen normally in PBF. 10.2 MYELOPOIESIS This is the process of formation of myeloid cells. It is restricted to the bone marrow after birth. The committed progenitor cell for granulocytes and monocytes is the GM-CFU which proliferates and differentiates to form myeloblast and monoblast. The myeloblast is the earliest morphologically identifiable cell. It is 10-18µm in size. The cytoplasm is scant and basophilic, usually agranular and may contain a few azurophilic cytoplasmic granules in the blast transiting to the next stage 80 HEMATOLOGY AND BLOOD BANK TECHNIQUE Maturation and Development of Leucocytes MODULE of promyelocyte. It has a large round to oval nucleus with a smooth nuclear Hematology and Blood membrane. The chromatin is fine, lacy and is evenly distributed throughout the Bank Technique nucleus. Two-five nucleoli can be identified in the nucleus. The next stage of maturation is the Promyelocyte. It is larger than a myeloblast, 12-20 µm with more abundant cytoplasm which has abundant primary azurophilic granules .