Identification and Characterization of the Monoblast in Mononuclear Phagocyte Colonies Grown in Vitro*
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Chapter 18 *Lecture Powerpoint the Circulatory System: Blood
Chapter 18 *Lecture PowerPoint The Circulatory System: Blood *See separate FlexArt PowerPoint slides for all figures and tables preinserted into PowerPoint without notes. Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Introduction • Many myths about blood – Mysterious ―vital force‖ – Drained ―bad-blood‖ for medical reasons – Hereditary traits were once thought to be transmitted through blood • Blood cells were seen with the first microscopes • Hematology—the study of blood • Recent developments in this field help save lives 18-2 Introduction • Expected Learning Outcomes – Describe the functions and major components of the circulatory system. – Describe the components and physical properties of blood. – Describe the composition of blood plasma. – Explain the significance of blood viscosity and osmolarity. – Describe in general terms how blood is produced. 18-3 Functions of the Circulatory System • Circulatory system consists of the heart, blood vessels, and blood • Cardiovascular system refers only to the heart and blood vessels • Hematology—the study of blood • Functions of circulatory system – Transport • O2, CO2, nutrients, wastes, hormones, and stem cells – Protection • Inflammation, limit spread of infection, destroy microorganisms and cancer cells, neutralize toxins, and initiate clotting – Regulation • Fluid balance, stabilizes pH of ECF, and temperature control 18-4 Components and General Properties of Blood • Adults have 4 to 6 L of blood • A liquid connective tissue consisting of -

Development of Plasmacytoid and Conventional Dendritic Cell Subtypes from Single Precursor Cells Derived in Vitro and in Vivo
ARTICLES Development of plasmacytoid and conventional dendritic cell subtypes from single precursor cells derived in vitro and in vivo Shalin H Naik1,2, Priyanka Sathe1,3, Hae-Young Park1,4, Donald Metcalf1, Anna I Proietto1,3, Aleksander Dakic1, Sebastian Carotta1, Meredith O’Keeffe1,4, Melanie Bahlo1, Anthony Papenfuss1, Jong-Young Kwak1,4,LiWu1 & Ken Shortman1 The development of functionally specialized subtypes of dendritic cells (DCs) can be modeled through the culture of bone marrow with the ligand for the cytokine receptor Flt3. Such cultures produce DCs resembling spleen plasmacytoid DCs (pDCs), http://www.nature.com/natureimmunology CD8+ conventional DCs (cDCs) and CD8– cDCs. Here we isolated two sequential DC-committed precursor cells from such cultures: dividing ‘pro-DCs’, which gave rise to transitional ‘pre-DCs’ en route to differentiating into the three distinct DC subtypes (pDCs, CD8+ cDCs and CD8– cDCs). We also isolated an in vivo equivalent of the DC-committed pro-DC precursor cell, which also gave rise to the three DC subtypes. Clonal analysis of the progeny of individual pro-DC precursors demonstrated that some pro-DC precursors gave rise to all three DC subtypes, some produced cDCs but not pDCs, and some were fully committed to a single DC subtype. Thus, commitment to particular DC subtypes begins mainly at this pro-DC stage. Dendritic cells (DCs) are antigen-presenting cells crucial for the innate macrophages12. Further ‘downstream’, ‘immediate’ precursors have and adaptive response to infection as well as for maintaining immune been identified for several DC types, including Ly6Chi monocytes as 3,4,6 13 Nature Publishing Group Group Nature Publishing tolerance to self tissue. -

The Biochemical and Biophysical Mechanisms of Macrophage Migration
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations 2015 The Biochemical and Biophysical Mechanisms of Macrophage Migration Laurel Erin Hind University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Allergy and Immunology Commons, Biomedical Commons, Immunology and Infectious Disease Commons, and the Medical Immunology Commons Recommended Citation Hind, Laurel Erin, "The Biochemical and Biophysical Mechanisms of Macrophage Migration" (2015). Publicly Accessible Penn Dissertations. 1062. https://repository.upenn.edu/edissertations/1062 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/1062 For more information, please contact [email protected]. The Biochemical and Biophysical Mechanisms of Macrophage Migration Abstract The ability of macrophages to migrate is critical for a proper immune response. During an innate immune response, macrophages migrate to sites of infection or inflammation where they clear pathogens through phagocytosis and activate an adaptive immune response by releasing cytokines and acting as antigen- presenting cells. Unfortunately, improper regulation of macrophage migration is associated with a variety of dieases including cancer, atherosclerosis, wound-healing, and rheumatoid arthritis. In this thesis, engineered substrates were used to study the chemical and physical mechanisms of macrophage migration. We first used microcontact printing to generate surfaces -

Of THP-1 Acute Monocytic Leukemia Cells Halt Proliferation and Induce
Human Plasma Membrane-Derived Vesicles Halt Proliferation and Induce Differentiation of THP-1 Acute Monocytic Leukemia Cells This information is current as Ephraim A. Ansa-Addo, Sigrun Lange, Dan Stratton, of September 28, 2021. Samuel Antwi-Baffour, Igor Cestari, Marcel I. Ramirez, Maria V. McCrossan and Jameel M. Inal J Immunol 2010; 185:5236-5246; Prepublished online 4 October 2010; doi: 10.4049/jimmunol.1001656 Downloaded from http://www.jimmunol.org/content/185/9/5236 References This article cites 43 articles, 12 of which you can access for free at: http://www.jimmunol.org/content/185/9/5236.full#ref-list-1 http://www.jimmunol.org/ Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists by guest on September 28, 2021 • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2010 by The American Association of Immunologists, Inc. All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology Human Plasma Membrane-Derived Vesicles Halt Proliferation and Induce Differentiation of THP-1 Acute Monocytic Leukemia Cells Ephraim A. -

Hematopoietic Stem-Cell Defects Underlying Abnormal Macrophage Development and Maturation in NOD/Lt Mice
Proc. Natl. Acad. Sci. USA Vol. 90, pp. 9625-9629, October 1993 Immunology Hematopoietic stem-cell defects underlying abnormal macrophage development and maturation in NOD/Lt mice: Defective regulation of cytokine receptors and protein kinase C DAVID V. SERREZE, JENS W. GAEDEKE, AND EDWARD H. LEITER* The Jackson Laboratory, 600 Main Street, Bar Harbor, ME 04609 Communicated by Elizabeth S. Russell, July 19, 1993 ABSTRACT The immunopathogenesis of autoimmune in- I-Ek, Db) of NON blocked the development of diabetogenic sulin-dependent diabetes in NOD mice entails defects in the T cells from NOD bone marrow (4). In addition, the inability development of macrophages (M$s) from hematopoietic pre- of NOD APCs to activate immunoregulatory T cells in a cursors. The present study analyzes the cellular and molecular syngeneic mixed-lymphocyte reaction was also found to be basis underlying our previous finding that the MO growth associated with homozygous expression of H-2g7 (5). factor colony-stimulating factor 1 (CSF-1) promotes a reduced Interactions between the H-2g7 haplotype and non-MHC- level of promonocyte proliferation and MO development from linked background genes also underlie the MO developmental NOD bone marrow. CSF-1 stimulation of NOD marrow in- anomalies characteristic of NOD mice. NOD bone marrow duced Mos to differentiate to the point that they secreted levels cells proliferate poorly in response to several myeloid growth of tumor necrosis factor a equivalent to that of controls. factors (6, 7). The myeloid growth factor, colony-stimulating However, CSF-1 failed to prime NOD M$s to completely factor 1 (CSF-1) generates fewer phenotypically mature differentiate in response to y-interferon, as shown by their (Mac-3+) M6s from NOD marrow than from diabetes- decreased lipopolysaccharide-stimulated interleukin 1 secre- resistant strains (6). -

White Blood Cells)
Lec.4 Medical Physiology – Blood Physiology Z.H.Kamil Leukocytes (White Blood Cells) Leukocytes are the only formed elements that are complete cells, with nuclei and the usual organelles. Accounting for less than 1% of total blood volume, leukocytes are far less numerous than red blood cells. On average, there are 4800–10,800 WBCs/μl of blood. Leukocytes are crucial to our defense against disease. They form a mobile army that helps protect the body from damage by bacteria, viruses, parasites, toxins, and tumor cells. As such, they have special functional characteristics. Red blood cells are kept into the bloodstream, and they carry out their functions in the blood. But white blood cells are able to slip out of the capillary blood vessels in a process called diapedesis and the circulatory system is simply their means of transport to areas of the body (mostly loose connective tissues or lymphoid tissues) where they mount inflammatory or immune responses. The signals that prompt WBCs to leave the bloodstream at specific locations are cell adhesion molecules displayed by endothelial cells forming the capillary walls at sites of inflammation. Once out of the bloodstream, leukocytes move through the tissue spaces by amoeboid motion (they form flowing cytoplasmic extensions that move them along). By following the chemical trail of molecules released by damaged cells or other leukocytes, a phenomenon called positive chemotaxis, they pinpoint areas of tissue damage and infection and gather there in large numbers to destroy foreign substances and dead cells. Whenever white blood cells are mobilized for action, the body speeds up their production and their numbers may double within a few hours. -

Download As A
QUANTITATIVE STUDY ON THE PRODUCTION AND KINETICS OF MONONUCLEAR PHAGOCYTES DURING AN ACUTE INFLAMMATORY REACTION BY RALPH VAN FURTH, MARTINA M. C. DIESSELHOFF-DEN DULK, AND HERMAN MATTIE* (From the Department of Microbial Diseases, University Hospital, Leiden, The Netherlands) (Received for publication 10 July 1973) During an acute inflammatory response in the peritoneal cavity both the peritoneal macrophages and the monocytes in the peripheral blood increase in number (1, 2). Labeling studies with [3H]thyrnidine have demonstrated that during an acute inflam- mation, just as during the normal steady state, the peritoneal macrophages derive from peripheral blood monocytes (1), which originate in the bone marrow from divid- ing promonocytes (3, 4). Although the mitotic activity of the promonocytes (DNA-synthesis time and cell cycle time) and the kinetic characteristics of these cells have been studied quantita- tively under steady-state conditions (4) and during treatment with glucocortico- steroids (5), similar data are not available for the acute inflammatory response. The present quantitative study, performed during an acute inflammation, provided data on the production of monocytes in the bone marrow, the influx and efflux of these cells into and from the peripheral blood, and the migration of monocytes into inflammatory lesion of the peritoneal cavity. These results were compared with the findings during the normal steady state. Materials and Methods Animals.--Specific pathogen-free male Swiss mice weighing 25-30 g (Central Institute for the Breeding of Laboratory Animals, TNO, Bilthoven, The Netherlands) were used. Cell Cultures.--The techniques for harvesting and culturing mouse bone marrow cells and peritoneal macrophages have been described in detail elsewhere (1, 3). -

Single-Cell Mass Cytometry of Differential Immune and Drug
RESEARCH ARTICLE Performance assessmentofmasscytometry. The workflow for mass cytometry is comparable with that of fluorescence flow cytometry (Fig. 1A). Single-Cell Mass Cytometry of Differential Antibodies coupled to distinct, stable, transition element isotopes were used to bind target epitopes on and within cells. Cells, with bound antibody- Immune and Drug Responses Across isotope conjugates, were sprayed as single-cell droplets into an inductively coupled argon plasma a Human Hematopoietic Continuum (created by passing argon gas through an induc- tion coil with a high radio-frequency electric cur- 1 1 2 3 1 1 Sean C. Bendall, * Erin F. Simonds, * Peng Qiu, El-ad D. Amir, Peter O. Krutzik, Rachel Finck, rent) at approximately 5500 K. This vaporizes 1,7 3 1 4,5 6 Robert V. Bruggner, Rachel Melamed, Angelica Trejo, Olga I. Ornatsky, Robert S. Balderas, each cell and induces ionization of its atomic con- 2 1 3 4,5 1 Sylvia K. Plevritis, Karen Sachs, Dana Pe’er, Scott D. Tanner, Garry P. Nolan † stituents. The resulting elemental ions were then sampled by a time-of-flight (TOF) mass spectrom- Flow cytometry is an essential tool for dissecting the functional complexity of hematopoiesis. We used eter and quantified. The signal for each transi- single-cell “mass cytometry” to examine healthy human bone marrow, measuring 34 parameters tion element isotope reporter was integrated as simultaneously in single cells (binding of 31 antibodies, viability, DNA content, and relative cell size). The each cell’s constituent ions reached the detector. signaling behavior of cell subsets spanning a defined hematopoietic hierarchy was monitored with 18 Currently, TOF sampling resolution enables the simultaneous markers of functional signaling states perturbed by a set of ex vivo stimuli and inhibitors. -

Platelet-Rich Plasma Regenerative Medicine: Sports Medicine, Orthopedic, and Recovery of Musculoskeletal Injuries Lecture Notes in Bioengineering
Lecture Notes in Bioengineering José Fábio Santos Duarte Lana Maria Helena Andrade Santana William Dias Belangero Angela Cristina Malheiros Luzo Editors Platelet-Rich Plasma Regenerative Medicine: Sports Medicine, Orthopedic, and Recovery of Musculoskeletal Injuries Lecture Notes in Bioengineering For further volumes: http://www.springer.com/series/11564 José Fábio Santos Duarte Lana Maria Helena Andrade Santana William Dias Belangero • Angela Cristina Malheiros Luzo Editors Platelet-Rich Plasma Regenerative Medicine: Sports Medicine, Orthopedic, and Recovery of Musculoskeletal Injuries 123 Editors José Fábio Santos Duarte Lana William Dias Belangero Research Institute of Sports Medicine Department of Orthopaedic and Orthopedics and Regeneration—iMOR Traumatology, Faculty of Medical Uberaba, MG Sciences Brazil University of Campinas Campinas, SP and Brazil Member of the Regenerative Angela Cristina Malheiros Luzo and Cell Therapy Group, Hemocentro Haematology and Hemotherapy Center University of Campinas, Unicamp Umbilical Cord Blood Bank Campinas, SP University of Campinas Brazil Campinas, SP Brazil Maria Helena Andrade Santana Department of Materials and Bioprocesses Engineering, School of Chemical Engineering University of Campinas Campinas, SP Brazil ISSN 2195-271X ISSN 2195-2728 (electronic) ISBN 978-3-642-40116-9 ISBN 978-3-642-40117-6 (eBook) DOI 10.1007/978-3-642-40117-6 Springer Heidelberg New York Dordrecht London Library of Congress Control Number: 2013950744 Ó Springer-Verlag Berlin Heidelberg 2014 This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. -

Monocyte and Macrophage Heterogeneity
REVIEWS MONOCYTE AND MACROPHAGE HETEROGENEITY Siamon Gordon and Philip R. Taylor Abstract | Heterogeneity of the macrophage lineage has long been recognized and, in part, is a result of the specialization of tissue macrophages in particular microenvironments. Circulating monocytes give rise to mature macrophages and are also heterogeneous themselves, although the physiological relevance of this is not completely understood. However, as we discuss here, recent studies have shown that monocyte heterogeneity is conserved in humans and mice, allowing dissection of its functional relevance: the different monocyte subsets seem to reflect developmental stages with distinct physiological roles, such as recruitment to inflammatory lesions or entry to normal tissues. These advances in our understanding have implications for the development of therapeutic strategies that are targeted to modify particular subpopulations of monocytes. OSTEOCLAST Circulating monocytes give rise to a variety of tissue- elicit increased recruitment of monocytes to peripheral A multinucleate cell that resident macrophages throughout the body, as well as sites4, where differentiation into macrophages and DCs resorbs bone. to specialized cells such as dendritic cells (DCs) and occurs, contributing to host defence, and tissue remod- OSTEOCLASTS. Monocytes are known to originate in the elling and repair. Studies of the mononuclear-phagocyte bone marrow from a common myeloid progenitor that system, using monoclonal antibodies specific for vari- is shared with neutrophils, and they are then released ous cell-surface receptors and differentiation antigens, into the peripheral blood, where they circulate for have shown that there is substantial heterogeneity of several days before entering tissues and replenishing phenotype, which most probably reflects the special- the tissue macrophage populations1. -
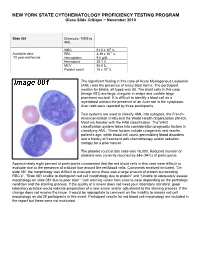
10 11 Cyto Slides 81-85
NEW YORK STATE CYTOHEMATOLOGY PROFICIENCY TESTING PROGRAM Glass Slide Critique ~ November 2010 Slide 081 Diagnosis: MDS to AML 9 WBC 51.0 x 10 /L 12 Available data: RBC 3.39 x 10 /L 72 year-old female Hemoglobin 9.6 g/dL Hematocrit 29.1 % MCV 86.0 fL Platelet count 16 x 109 /L The significant finding in this case of Acute Myelogenous Leukemia (AML) was the presence of many blast forms. The participant median for blasts, all types was 88. The blast cells in this case (Image 081) are large, irregular in shape and contain large prominent nucleoli. It is difficult to identify a blast cell as a myeloblast without the presence of an Auer rod in the cytoplasm. Auer rods were reported by three participants. Two systems are used to classify AML into subtypes, the French- American-British (FAB) and the World Health Organization (WHO). Most are familiar with the FAB classification. The WHO classification system takes into consideration prognostic factors in classifying AML. These factors include cytogenetic test results, patient’s age, white blood cell count, pre-existing blood disorders and a history of treatment with chemotherapy and/or radiation therapy for a prior cancer. The platelet count in this case was 16,000. Reduced number of platelets was correctly reported by 346 (94%) of participants. Approximately eight percent of participants commented that the red blood cells in this case were difficult to evaluate due to the presence of a bluish hue around the red blood cells. Comments received included, “On slide 081 the morphology was difficult to evaluate since there was a large amount of protein surrounding RBC’s”, “Slide 081 unable to distinguish red cell morphology due to protein” and “Unable to adequately assess morphology on slide 081 due to poor stain”. -

10 Maturation and Development of Leucocytes
MODULE Maturation and Development of Leucocytes Hematology and Blood Bank Technique 10 Notes MATURATION AND DEVELOPMENT OF LEUCOCYTES 10.1 INTRODUCTION The leucocytes develop from the multipotent hematopoietic stem cell which then gives rise to a stem cell committed to formation of leucocytes. Both these cells cannot be identified morphologically by routine methods. The various types of leucocytes are granulocytes (neutrophils, eosinophils and basophils), monocytes and lymphocytes. The three cell types develop separately and accordingly these processes will be discussed separately. OBJECTIVES After reading this lesson, you will be able to: z explain the various stages in the development of leucocytes. z describe the different types of leucocytes seen normally in PBF. 10.2 MYELOPOIESIS This is the process of formation of myeloid cells. It is restricted to the bone marrow after birth. The committed progenitor cell for granulocytes and monocytes is the GM-CFU which proliferates and differentiates to form myeloblast and monoblast. The myeloblast is the earliest morphologically identifiable cell. It is 10-18µm in size. The cytoplasm is scant and basophilic, usually agranular and may contain a few azurophilic cytoplasmic granules in the blast transiting to the next stage 80 HEMATOLOGY AND BLOOD BANK TECHNIQUE Maturation and Development of Leucocytes MODULE of promyelocyte. It has a large round to oval nucleus with a smooth nuclear Hematology and Blood membrane. The chromatin is fine, lacy and is evenly distributed throughout the Bank Technique nucleus. Two-five nucleoli can be identified in the nucleus. The next stage of maturation is the Promyelocyte. It is larger than a myeloblast, 12-20 µm with more abundant cytoplasm which has abundant primary azurophilic granules .