An Ecological Valence Theory of Human Color Preference
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

COLOR SPACE MODELS for VIDEO and CHROMA SUBSAMPLING
COLOR SPACE MODELS for VIDEO and CHROMA SUBSAMPLING Color space A color model is an abstract mathematical model describing the way colors can be represented as tuples of numbers, typically as three or four values or color components (e.g. RGB and CMYK are color models). However, a color model with no associated mapping function to an absolute color space is a more or less arbitrary color system with little connection to the requirements of any given application. Adding a certain mapping function between the color model and a certain reference color space results in a definite "footprint" within the reference color space. This "footprint" is known as a gamut, and, in combination with the color model, defines a new color space. For example, Adobe RGB and sRGB are two different absolute color spaces, both based on the RGB model. In the most generic sense of the definition above, color spaces can be defined without the use of a color model. These spaces, such as Pantone, are in effect a given set of names or numbers which are defined by the existence of a corresponding set of physical color swatches. This article focuses on the mathematical model concept. Understanding the concept Most people have heard that a wide range of colors can be created by the primary colors red, blue, and yellow, if working with paints. Those colors then define a color space. We can specify the amount of red color as the X axis, the amount of blue as the Y axis, and the amount of yellow as the Z axis, giving us a three-dimensional space, wherein every possible color has a unique position. -

Tabla De Conversión Pantone a NCS (Natural Color System)
Tabla de conversión Pantone a NCS (Natural Color System) PANTONE NCS (más parecido) PANTONE NCS (más parecido) Pantone Yellow C NCS 0580-Y Pantone 3985C NCS 3060-G80Y Pantone Yellow U NCS 0580-Y Pantone 3985U NCS 4040-G80Y Pantone Warm Red C NCS 0580-Y70R Pantone 3995C NCS 5040-G80Y Pantone Warm Red U NCS 0580-Y70R Pantone 3995U NCS 6020-G70Y Pantone Rubine Red C NCS 1575-R10B Pantone 400C NCS 2005-Y50R Pantone Rubine Red U NCS 1070-R20B Pantone 400U NCS 2502-R Pantone Rhodamine Red C Pantone 401C NCS 2005-Y50R Pantone Rhodamine Red U NCS 1070-R20B Pantone 401U NCS 2502-R Pantone Purple C Pantone 402C NCS 4005-Y50R Pantone Purple U NCS 2060-R40B Pantone 402U NCS 3502-R Pantone Violet C Pantone 403C NCS 4055-Y50R Pantone Violet U NCS 3050-R60B Pantone 403U NCS 4502-R Pantone Reflex Blue C NCS 3560-R80B Pantone 404C NCS 6005-Y20R Pantone Reflex Blue U NCS 3060-R70B Pantone 404U NCS 5502-R Pantone Process Blue C NCS 2065-B Pantone 405C NCS 7005-Y20R Pantone Process Blue U NCS 1565-B Pantone 405U NCS 6502-R Pantone Green C NCS 2060-B90G Pantone 406C NCS 2005-Y50R Pantone Green U NCS 2060-B90G Pantone 406U NCS 2005-Y50R Pantone Black C NCS 8005-Y20R Pantone 407C NCS 3005-Y50R Pantone Black U NCS 7502-Y Pantone 407U NCS 3005-Y80R Pantone Yellow 012C NCS 0580-Y Pantone 408C NCS 3005-Y50R Pantone Yellow 012U NCS 0580-Y Pantone 408U NCS 4005-Y80R Pantone Orange 021C NCS 0585-Y60R Pantone 409C NCS 5005-Y50R Pantone Orange 021U NCS 0580-Y60R Pantone 409U NCS 5005-Y80R Pantone Red 032C NCS 0580-Y90R Pantone 410C NCS 5005-Y50R Pantone Red 032U NCS -

RAINSTONE Inspired by Nature, This Collection Will Give Fresh and Natural Design Elements to Any Space
RAINSTONE Inspired by nature, this collection will give fresh and natural design elements to any space. Rain Stone Dark Grey 60x60 cm | 24”x 24” Dark Grey Rain Stone 12 13 Floor Floor Floor Rain Stone_Natural 60x60 | 24”x24” 14 15 RAIN STONE NATURAL RAIN STONE BEIGE WHITE *CERST060002_NATURAL 60x120 - 24”x48” rect. CERST060006_NATURAL 60x60 - 24”x24” rect. *CERST060001_BEIGE WHITE 60x120 - 24”x48” rect. CERST060005_BEIGE WHITE 60x60 - 24”x24” rect. *CERST030014 _NATURAL 30x120 - 12”x48” rect. CERST030002_NATURAL 30x60 - 12”x24” rect. *CERST030013 _BEIGE WHITE 30x120 - 12”x48” rect. CERST030001_BEIGE WHITE 30x60 - 12”x24” rect. *CERST029002_NATURAL 29x29 - 12”x12” rect. CERST005002_NATURAL MOSAICO *CERST029001_BEIGE WHITE 29x29 - 12”x12” rect. CERST005001_BEIGE WHITE MOSAICO 5x5 - 2”x2” rect. 5x5 - 2”x2” rect. *CERST015002_NATURAL 15x60 - 6”x24” rect. *CERST010002_NATURAL 10x30 - 4”x12” rect. *CERST015001_BEIGE WHITE 15x60 - 6”x24” rect. *CERST010001_BEIGE WHITE 10x30 - 4”x12” rect. *CERST030006_NATURAL *CERST030010_NATURAL WALL *CERST030005_BEIGE WHITE *CERST030009_ BEIGE WHITE WALL 5x15 - 2”x6” rect. 30x60 - 12”x24” rect. 5x15 - 2”x6” rect. 30x60 - 12”x24” rect. 30x30 - 12”x12” su rete rect. 30x30 - 12”x12” su rete rect. *CERST031002 _ NATURAL CHEVRON *CERST028002 _ NATURAL FRINGE *CERST031001_ BEIGE WHITE CHEVRON *CERST028001_ BEIGE WHITE FRINGE 31,5x29,6 - 12,40”x11,65” rect. 28,8X28,8 - 11,34”x11,34” rect. 31,5x29,6 - 12,40”x11,65” rect. 28,8X28,8 - 11,34”x11,34” rect. 16 * Special order sizes 17 RAIN STONE LIGHT GREY RAIN STONE DARK GREY *CERST060003_LIGHT GREY 60x120 - 24”x48” rect. CERST060007_LIGHT GREY 60x60 - 24”x24” rect. *CERST060004_DARK GREY 60x120 - 24”x48” rect. CERST060008_DARK GREY 60x60 - 24”x24” rect. -
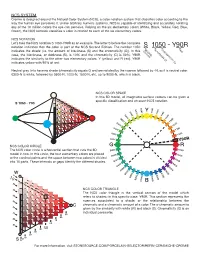
Cromie Is Designed Around the Natural Color System (NCS), a Color Notation System That Classifies Color According to the Way the Human Eye Perceives It
NCS SYSTEM Cromie is designed around the Natural Color System (NCS), a color notation system that classifies color according to the way the human eye perceives it. Unlike arbitrary numeric systems, NCS is capable of identifying and accurately notating any of the 10 million colors the eye can perceive. Relying on the six elementary colors (White, Black, Yellow, Red, Blue, Green), the NCS notation classifies a color in relation to each of the six elementary colors. NCS NOTATION Let’s take the NCS notation S 1050-Y90R as an example. The letter S before the complete notation indicates that the color is part of the NCS Second Edition. The number 1050 indicates the shade (i.e. the amount of blackness (S) and the chromaticity (C)). In this case, the blackness, or darkness (S), is 10% and the chromaticity (C) is 50%. Y90R indicates the similarity to the other two elementary colors, Y (yellow) and R (red). Y90R indicates yellow with 90% of red. Neutral gray tints have no shade (chromaticity equals 0) and are notated by the nuance followed by -N, as it is neutral color. 0300-N is white, followed by 0500-N, 1000-N, 1500 N, etc. up to 9000-N, which is black. NCS COLOR SPACE In this 3D model, all imaginable surface colours can be given a specific classification and an exact NCS notation. S 1050 - Y90 NCS COLOR CIRCLE The NCS color circle is a horizontal section that cuts the 3D model in two. In this circle, the four elementary colors are placed at the cardinal points and the space between two colors is divided into 10 parts. -

Color Appearance Models Second Edition
Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Color Appearance Models Wiley–IS&T Series in Imaging Science and Technology Series Editor: Michael A. Kriss Formerly of the Eastman Kodak Research Laboratories and the University of Rochester The Reproduction of Colour (6th Edition) R. W. G. Hunt Color Appearance Models (2nd Edition) Mark D. Fairchild Published in Association with the Society for Imaging Science and Technology Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Copyright © 2005 John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England Telephone (+44) 1243 779777 This book was previously publisher by Pearson Education, Inc Email (for orders and customer service enquiries): [email protected] Visit our Home Page on www.wileyeurope.com or www.wiley.com All Rights Reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, scanning or otherwise, except under the terms of the Copyright, Designs and Patents Act 1988 or under the terms of a licence issued by the Copyright Licensing Agency Ltd, 90 Tottenham Court Road, London W1T 4LP, UK, without the permission in writing of the Publisher. Requests to the Publisher should be addressed to the Permissions Department, John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England, or emailed to [email protected], or faxed to (+44) 1243 770571. This publication is designed to offer Authors the opportunity to publish accurate and authoritative information in regard to the subject matter covered. -

Major Color Order Systems and Their Psychophysical Structure
Chapter 7 Major Color Order Systems and Their Psychophysical Structure In this chapter only the Munsell, OSA-UCS, and Swedish NCS systems will be discussed. The former two are the most important attempts to create psy- chologically uniform systems, the latter uses the presumed natural approach of Hering (see Chapter 2) to create a color order system, having a regular structure, but not one uniform in terms of size of perceived differences. There are several other newer color order systems extant, but they neither make claims for uniformity nor for regularity according to new, significant psycho- logical attributes. The issue of viewing conditions for these systems has been attended to in different ways.As discussed previously,the Munsell system is illustrated as if the chips at each value level would be viewed against an achromatic surround of the same value.Chips of two adjacent value levels are illustrated as if viewed against an achromatic surround of intermediate value. The actual atlas displays the chips on white paper (historically of various degrees of whiteness), thus result- ing in distortions of the value scale, particularly at lower values.The OSA-UCS system is defined for an achromatic surround of luminous reflectance Y = 30 (L = 0).The atlas samples are in transparent jackets.NCS,finally,has been estab- lished against an immediate achromatic surround of Y = 78 in a light booth painted with an achromatic gray of Y = 54.The atlas displays samples on white paper. Both latter systems only result in the intended color experiences when viewed against the appropriate surrounds. Munsell and NCS are defined for Color Space and Its Divisions: Color Order from Antiquity to the Present, by Rolf G. -

Tfl Basic Elements Standard
Transport for London Transport for London Basic elements standard Issue 3 MAYOR OF LONDON Transport for London Contents Foreword 1 Glossary 1.1 Colour glossary 1.2 Corporate typeface 2 Transport for London 2.1 Roundel usage 2.2 Roundel usage continued 2.3 Exclusion zones 2.4 Colours 3 Secondary identities For further information Foreword Contents A well designed, confident and consistent visual identity is highly effective in communicating the strengths of our organisation. It is essential that Transport for London (TfL) maintains a high standard for co-ordinated design in every aspect of our operations. This document gives guidance on the basic elements that make up the TfL visual identity. The information covers the corporate typeface, the visual identity and the corporate colours. For more guidance on TfL Corporate standards, please visit the TfL website: tfl.gov.uk/corporatedesign 1 Glossary Contents The main terms used in these guidelines are Symbol Logotype as follows: Other visual identifiers used by TfL. The text of TfL set in New Johnston Medium upper and lower case type. Logotypes are Mark used with roundels or symbols in a fixed A combination of a roundel or symbol relationship to form Marks, but can also be with a logotype in a fixed relationship as used in a looser relationship such as that used shown below. on stationery (see the Stationery standards). Exclusion zone The minimum area around marks, roundels or Transport for London symbols that must be kept free of other graphic elements. This is to ensure that they are reproduced clearly and legibly without Roundel interference from other visual devices. -

Color Appearance Models Second Edition
Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratoiy Rochester Institute of Technology, USA John Wiley & Sons, Ltd Contents Series Preface xiii Preface XV Introduction xix 1 Human Color Vision 1 1.1 Optics of the Eye 1 1.2 The Retina 6 1.3 Visual Signal Processing 12 1.4 Mechanisms of Color Vision 17 1.5 Spatial and Temporal Properties of Color Vision 26 1.6 Color Vision Deficiencies 30 1.7 Key Features for Color Appearance Modeling 34 2 Psychophysics 35 2.1 Psychophysics Defined 36 2.2 Historical Context 37 2.3 Hierarchy of Scales 40 2.4 Threshold Techniques 42 2.5 Matching Techniques 45 2.6 One-Dimensional Scaling 46 2.7 Multidimensional Scaling 49 2.8 Design of Psychophysical Experiments 50 2.9 Importance in Color Appearance Modeling 52 3 Colorimetry 53 3.1 Basic and Advanced Colorimetry 53 3.2 Whyis Color? 54 3.3 Light Sources and Illuminants 55 3.4 Colored Materials 59 3.5 The Human Visual Response 66 3.6 Tristimulus Values and Color Matching Functions 70 3.7 Chromaticity Diagrams 77 3.8 CIE Color Spaces 78 3.9 Color Difference Specification 80 3.10 The Next Step , 82 viii CONTENTS 4 Color Appearance Terminology 83 4.1 Importance of Definitions 83 4.2 Color 84 4.3 Hue 85 4.4 Brightness and Lightness 86 4.5 Colorfulness and Chroma 87 4.6 Saturation 88 4.7 Unrelated and Related Colors 88 4.8 Definitions in Equations 90 4.9 Brightness-Colorfulness vs Lightness-Chroma 91 5 Color Order Systems 94 5.1 Overvlew and Requirements 94 5.2 The Munsell Book of Color 96 5.3 The Swedish Natural Color System (NCS) -

Color Appearance and Color Difference Specification
Errata: 1) On page 200, the coefficient on B^1/3 for the j coordinate in Eq. 5.2 should be -9.7, rather than the +9.7 that is published. 2) On page 203, Eq. 5.4, the upper expression for L* should be L* = 116(Y/Yn)^1/3-16. The exponent 1/3 is omitted in the published text. In addition, the leading factor in the expression for b* should be 200 rather than 500. 3) In the expression for "deltaH*_94" just above Eq. 5.8, each term inside the radical should be squared. Color Appearance and Color Difference Specification David H. Brainard Department of Psychology University of California, Santa Barbara, CA 93106, USA Present address: Department of Psychology University of Pennsylvania, 381 S Walnut Street Philadelphia, PA 19104-6196, USA 5.1 Introduction 192 5.3.1.2 Definition of CI ELAB 202 5.31.3 Underlying experimental data 203 5.2 Color order systems 192 5.3.1.4 Discussion olthe C1ELAB system 203 5.2.1 Example: Munsell color order system 192 5.3.2 Other color difference systems 206 5.2.1.1 Problem - specifying the appearance 5.3.2.1 C1ELUV 206 of surfaces 192 5.3.2.2 Color order systems 206 5.2.1.2 Perceptual ideas 193 5.2.1.3 Geometric representation 193 5.4 Current directions in color specification 206 5.2.1.4 Relating Munsell notations to stimuli 195 5.4.1 Context effects 206 5.2.1.5 Discussion 196 5.4.1.1 Color appearance models 209 5.2.16 Relation to tristimulus coordinates 197 5.4.1.2 C1ECAM97s 209 5.2.2 Other color order systems 198 5.4.1.3 Discussion 210 5.2.2.1 Swedish Natural Colour System 5.42 Metamerism 211 (NC5) 198 5.4.2.1 The -

Color for Philosophers: Unweaving the Rainbow
c o N T E N T 5 Foreword by Arthur Danto ix xv Preface xix Introduction I Color Perception and Science The physical causes of color 1 The camera and the eye 7 Perceiving lightness and darkness 19 26 Chromatic vision Chromatic response 36 The structure of phenomenal hues 40 Object metamerism, adaptation, and contrast 45 Some mechanisms of chromatic perception 52 II The Ontology of Color Objectivism 59 Standard conditions 67 Normal observers 76 Constancy and crudity 82 Ch romatic democracy 91 Sense data as color bearers 96 Materialist reduction and the illusion of color 109 III Phenomenology and Physiology THE RELATlONS OF COLORS TO EACH OTHER 113 T he resemblances of colors 113 The incompatibilities of colors 121 Deeper problems 127 OTHER MINDS 134 Spectral inversions and asymmetries 134 vii CONTENTS I nternalism and externalism 142 Other colors, other minds 145 COLOR LANGUAGE 155 Foci 155 The evolution of color categories 165 Boundaries and indeterminacy 169 Establishing boundaries 182 Color Plates following page 88 Appendix: Land's Retinex Theory of Color Vision 187 Notes 195 Glossary of Technical Terms 209 Further Reading 216 Bibliography 217 Acknowledgments 234 Indexes 237 viii F o R E w o R D Very few today still believe that philosophy is a disease of language and that its deliverances, due to disturbances of the grammatical un conscious, are neither true nor false but nonsense. But the fact re mains that, very often, philosophical theory stands to positive knowledge roughly in the relationship in which hysteria is said to stand to anatomical truth. -

COLOR APPEARANCE MODELS, 2Nd Ed. Table of Contents
COLOR APPEARANCE MODELS, 2nd Ed. Mark D. Fairchild Table of Contents Dedication Table of Contents Preface Introduction Chapter 1 Human Color Vision 1.1 Optics of the Eye 1.2 The Retina 1.3 Visual Signal Processing 1.4 Mechanisms of Color Vision 1.5 Spatial and Temporal Properties of Color Vision 1.6 Color Vision Deficiencies 1.7 Key Features for Color Appearance Modeling Chapter 2 Psychophysics 2.1 Definition of Psychophysics 2.2 Historical Context 2.3 Hierarchy of Scales 2.4 Threshold Techniques 2.5 Matching Techniques 2.6 One-Dimensional Scaling 2.7 Multidimensional Scaling 2.8 Design of Psychophysical Experiments 2.9 Importance in Color Appearance Modeling Chapter 3 Colorimetry 3.1 Basic and Advanced Colorimetry 3.2 Why is Color? 3.3 Light Sources and Illuminants 3.4 Colored Materials 3.5 The Visual Response 3.6 Tristimulus Values and Color-Matching Functions 3.7 Chromaticity Diagrams 3.8 CIE Color Spaces 3.9 Color-Difference Specification 3.10 The Next Step Chapter 4 Color Appearance Models: Table of Contents Page 1 Color-Appearance Terminology 4.1 Importance of Definitions 4.2 Color 4.3 Hue 4.4 Brightness and Lightness 4.5 Colorfulness and Chroma 4.6 Saturation 4.7 Unrelated and Related Colors 4.8 Definitions in Equations 4.9 Brightness-Colorfulness vs. Lightness-Chroma Chapter 5 Color Order Systems 5.1 Overview and Requirements 5.2 The Munsell Book of Color 5.3 The Swedish Natural Color System (NCS) 5.4 The Colorcurve System 5.5 Other Color Order Systems 5.6 Uses of Color Order Systems 5.7 Color Naming Systems Chapter 6 Color-Appearance -

Color Marketing Group 2006 Consumer Color Directions® Notations Chart
Color Marketing Group 2006 Consumer Color Directions® Notations Chart KEY 1 Munsell Asian Rose Bliss 2 Pantone Textile 1 5R 5/10 1 5YR 9/2 Pantone Matching System, 3 2 18-1648 2 12-1011 Coated/Uncoated 3 187C/1805U 3 475C/475U 4 Pantone Process Number 4 DS88-1C 4 DS36-8C 5 RAL 5 030 50 50 5 060 85 20 6 RGB 6 R-197 G-73 B-65 6 R-247 G-204 B-159 7 CMYK 7 C-0, M-100, Y-90, K-20 7 C-0, M-15, Y-25, K-0 8 NCS (Natural Color System) 8 S 2060-R 8 S 1010-Y50R 9 15-12-20 9 13-04-08 9 Archroma Burlesque Chillin' Cybernatural Diamante Silver 1 5R 3/6 1 10B 7/8 1 7.5Y 8.5/4 1 5PB 8/0.5 2 19-1338 2 14-4516 2 13-0633 2 14-5002 TPX 3 505C/188U 3 7458C/630U 3 460C/461U 3 423C/877U 4 DS322-4C 4 DS232-6C 4 DS309-8C 4 DS325-6C 5 020 20 29 5 230 70 30 5 095 80 30 5 * 6 R-83 G-14 B-15 6 R-147 G-202 B-211 6 R-237 G-217 B-140 6 R-193 G-185 B-164 7 C-60, M-100, Y-100, K-0 7 C-40, M-0, Y-0, K-0 7 C-3, M-0, Y-30, K-0 7 C-0, M-0M Y-0, K-40 8 S 6030-R 8 S 0540-B 8 S 1020-G90Y 8 S 1005-R90B 9 15-14-06 9 26-02-08 9 21-08-28 9 22-04-02 Decoesque Electric Mud Elemental Gray Flemish Gold 1 7.5G 4/4 1 2.5R 2.5/2 1 5R 4/1 1 2.5Y 6/6 2 18-5622 2 19-1619 2 19-3803 2 17-1047 3 625C/555U 3 5185C/5185U 3 405C/7519U 3 1255C/1245U 4 DS269-4C 4 DS323-1C 4 DS326-3C 4 DS27-5C 5 160 50 20 5 020 20 05 5 060 40 05 5 070 60 40 6 R-86 G-126 B-92 6 R-75 G-53 B-50 6 R-104 G-94 B-80 6 R-180 G-133 B-50 7 C-60, M-0, Y-50, K-30 7 C-90, M-100, Y-100, K-15 7 C-70, M-55, Y-55, K-0 7 C-10, M-30, Y-70, K-10 8 S 5020-B90G 8 S 8010-R30B 8 S 6502-R 8 S 4050-Y10R 9 23-12-12 9 18-16-05 9 19-12-02 9 13-08-10