Ketone Body Metabolism Preserves Hepatic Function During Adaptation to Birth and in Overnutrition David Graham Cotter Washington University in St
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Turkish Patient with Succinyl-Coa:3-Oxoacid Coa
Original Article Journal of Inborn Errors of Metabolism & Screening 2016, Volume 4: 1–5 A Turkish Patient With ª The Author(s) 2016 DOI: 10.1177/2326409816651281 Succinyl-CoA:3-Oxoacid CoA iem.sagepub.com Transferase Deficiency Mimicking Diabetic Ketoacidosis Sahin Erdol, MD1, Mehmet Ture, MD2, Tahsin Yakut, PhD2, Halil Saglam, PhD1, Hideo Sasai, MD3, Elsayed Abdelkreem, MD3, Hiroki Otsuka, MD3, and Toshiyuki Fukao, MD, PhD3 Abstract Succinyl-CoA:3-oxoacid CoA transferase (SCOT) deficiency is an autosomal recessive disorder of ketone body utilization that is clinically characterized with intermittent ketoacidosis crises. We report here the second Turkish case with SCOT deficiency. She experienced 3 ketoacidotic episodes: The first ketoacidotic crisis mimicked diabetic ketoacidosis because of the associated hyperglycemia. Among patients with SCOT deficiency, the blood glucose levels at the first crises were variable, and this case had the highest ever reported blood glucose level. She is a compound heterozygote with 2 novel mutations, c.517A>G (K173E) and c.1543A>G (M515V), in exons 5 and 17 of the OXCT1 gene, respectively. In patient’s fibroblasts, SCOT activity was deficient and, by immunoblot analysis, SCOT protein was much reduced. The patient attained normal development and had no permanent ketosis. The accurate diagnosis of SCOT deficiency in this case had a vital impact on the management strategy and outcome. Keywords succinyl-CoA:3-oxoacid CoA transferase, deficiency, mimicking, diabetic ketoacidosis, Turkey Introduction deficiency. We herein describe a Turkish SCOT-deficient patient whose first episode mimicked diabetic ketoacidosis Succinyl-CoA:3-oxoacid CoA transferase (SCOT) deficiency because of the associated hyperglycemia. -

Generated by SRI International Pathway Tools Version 25.0, Authors S
An online version of this diagram is available at BioCyc.org. Biosynthetic pathways are positioned in the left of the cytoplasm, degradative pathways on the right, and reactions not assigned to any pathway are in the far right of the cytoplasm. Transporters and membrane proteins are shown on the membrane. Periplasmic (where appropriate) and extracellular reactions and proteins may also be shown. Pathways are colored according to their cellular function. Gcf_000238675-HmpCyc: Bacillus smithii 7_3_47FAA Cellular Overview Connections between pathways are omitted for legibility. -

Novel Compound Heterozygous OXCT1 Mutations Causing Succinyl-Coa:3-Ketoacid Coa Transferase Deficiency
Case Report Yonsei Med J 2019 Mar;60(3):308-311 https://doi.org/10.3349/ymj.2019.60.3.308 pISSN: 0513-5796 · eISSN: 1976-2437 A Rare Cause of Life-Threatening Ketoacidosis: Novel Compound Heterozygous OXCT1 Mutations Causing Succinyl-CoA:3-Ketoacid CoA Transferase Deficiency Young A Kim1,2, Seong Heon Kim1, Chong Kun Cheon1, and Yoo-Mi Kim3 1Department of Pediatrics, Pusan National University Children’s Hospital, Yangsan; 2Research Institute for Convergence of Biomedical Science and Technology, Pusan National University Yangsan Hospital, Yangsan; 3Department of Pediatrics, Chungnam National University Hospital, College of Medicine, Chungnam National University, Daejeon, Korea. Succinyl-CoA:3-ketoacid CoA transferase (SCOT) deficiency is a rare inborn error of ketone body utilization, characterized by ep- isodic or permanent ketosis. SCOT deficiency is caused by mutations in the OXCT1 gene, which is mapped to 5p13 and consists of 17 exons. A 12-month-old girl presented with severe ketoacidosis and was treated with continuous renal replacement therapy. She had two previously unrecognized mild-form episodes of ketoacidosis followed by febrile illness. While high levels of ketone bodies were found in her blood and urine, other laboratory investigations, including serum glucose, were unremarkable. We identified novel compound heterozygous mutations in OXCT1:c.1118T>G (p.Ile373Ser) and a large deletion ranging from exon 8 to 16 through targeted exome sequencing and microarray analysis. This is the first Korean case of SCOT deficiency caused by novel mutations in OXCT1, resulting in life-threatening ketoacidosis. In patients with unexplained episodic ketosis, or high anion gap metabolic acidosis in infancy, an inherited disorder in ketone body metabolism should be suspected. -

Yeast Genome Gazetteer P35-65
gazetteer Metabolism 35 tRNA modification mitochondrial transport amino-acid metabolism other tRNA-transcription activities vesicular transport (Golgi network, etc.) nitrogen and sulphur metabolism mRNA synthesis peroxisomal transport nucleotide metabolism mRNA processing (splicing) vacuolar transport phosphate metabolism mRNA processing (5’-end, 3’-end processing extracellular transport carbohydrate metabolism and mRNA degradation) cellular import lipid, fatty-acid and sterol metabolism other mRNA-transcription activities other intracellular-transport activities biosynthesis of vitamins, cofactors and RNA transport prosthetic groups other transcription activities Cellular organization and biogenesis 54 ionic homeostasis organization and biogenesis of cell wall and Protein synthesis 48 plasma membrane Energy 40 ribosomal proteins organization and biogenesis of glycolysis translation (initiation,elongation and cytoskeleton gluconeogenesis termination) organization and biogenesis of endoplasmic pentose-phosphate pathway translational control reticulum and Golgi tricarboxylic-acid pathway tRNA synthetases organization and biogenesis of chromosome respiration other protein-synthesis activities structure fermentation mitochondrial organization and biogenesis metabolism of energy reserves (glycogen Protein destination 49 peroxisomal organization and biogenesis and trehalose) protein folding and stabilization endosomal organization and biogenesis other energy-generation activities protein targeting, sorting and translocation vacuolar and lysosomal -
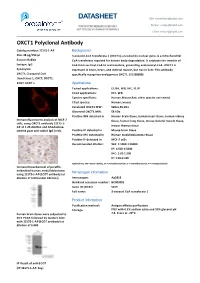
DATASHEET USA: [email protected]
DATASHEET USA: [email protected] FOR IN VITRO RESEARCH USE ONLY Europe: [email protected] NOT FOR USE IN HUMANS OR ANIMALS China: [email protected] OXCT1 Polyclonal Anbody Catalog number: 12175-1-AP Background Size: 38 μg/150 μl 3-oxoacid-CoA transferase 1 (OXCT1), encoded by nuclear gene, is a mitochondrial Source: Rabbit CoA transferase required for ketone body degradaon. It catalyzes the transfer of Isotype: IgG CoA from succinyl-CoA to acetoacetate, generang acetoacetyl-CoA. OXCT1 is Synonyms: expressed in brain, heart, and skeletal muscle, but not in liver. This anbody OXCT1; 3 oxoacid CoA specifically recognizes endogenous OXCT1. (21209089) transferase 1, OXCT, OXCT1, SCOT, SCOT s Applications Tested applicaons: ELISA, WB, IHC, IF, IP Cited applicaons: IHC, WB Species specificity: Human,Mouse,Rat; other species not tested. Cited species: Human, mouse Caculated OXCT1 MW: 520aa,56 kDa Observed OXCT1 MW: 56 kDa Posive WB detected in Human brain ssue, human heart ssue, human kidney Immunofluorescent analysis of MCF-7 ssue, human lung ssue, mouse skeletal muscle ssue, cells, using OXCT1 anbody 12175-1- AP at 1:25 diluon and Rhodamine- mouse thymus ssue labeled goat an-rabbit IgG (red). Posive IP detected in Mouse brain ssue Posive IHC detected in Human medulloblastoma ssue Posive IF detected in MCF-7 cells Recommended diluon: WB: 1:1000-1:10000 IP: 1:500-1:5000 IHC: 1:20-1:200 IF: 1:10-1:100 Applicaon key: WB = Western blong, IHC = Immunohistochemistry, IF = Immunofluorescence, IP = Immunoprecipitaon Immunohistochemical of paraffin- embedded human medulloblastoma Immunogen information using 12175-1-AP(SCOT anbody) at diluon of 1:50 (under 10x lens) Immunogen: Ag2818 GenBank accession number: BC009001 Gene ID (NCBI): 5019 Full name: 3-oxoacid CoA transferase 1 Product information Purificaon method: Angen affinity purificaon Storage: PBS with 0.1% sodium azide and 50% glycerol pH o human brain ssue were subjected to 7.3. -

Letters to Nature
letters to nature Received 7 July; accepted 21 September 1998. 26. Tronrud, D. E. Conjugate-direction minimization: an improved method for the re®nement of macromolecules. Acta Crystallogr. A 48, 912±916 (1992). 1. Dalbey, R. E., Lively, M. O., Bron, S. & van Dijl, J. M. The chemistry and enzymology of the type 1 27. Wolfe, P. B., Wickner, W. & Goodman, J. M. Sequence of the leader peptidase gene of Escherichia coli signal peptidases. Protein Sci. 6, 1129±1138 (1997). and the orientation of leader peptidase in the bacterial envelope. J. Biol. Chem. 258, 12073±12080 2. Kuo, D. W. et al. Escherichia coli leader peptidase: production of an active form lacking a requirement (1983). for detergent and development of peptide substrates. Arch. Biochem. Biophys. 303, 274±280 (1993). 28. Kraulis, P.G. Molscript: a program to produce both detailed and schematic plots of protein structures. 3. Tschantz, W. R. et al. Characterization of a soluble, catalytically active form of Escherichia coli leader J. Appl. Crystallogr. 24, 946±950 (1991). peptidase: requirement of detergent or phospholipid for optimal activity. Biochemistry 34, 3935±3941 29. Nicholls, A., Sharp, K. A. & Honig, B. Protein folding and association: insights from the interfacial and (1995). the thermodynamic properties of hydrocarbons. Proteins Struct. Funct. Genet. 11, 281±296 (1991). 4. Allsop, A. E. et al.inAnti-Infectives, Recent Advances in Chemistry and Structure-Activity Relationships 30. Meritt, E. A. & Bacon, D. J. Raster3D: photorealistic molecular graphics. Methods Enzymol. 277, 505± (eds Bently, P. H. & O'Hanlon, P. J.) 61±72 (R. Soc. Chem., Cambridge, 1997). -

Product Sheet Info
Master Clone List for NR-19279 ® Vibrio cholerae Gateway Clone Set, Recombinant in Escherichia coli, Plates 1-46 Catalog No. NR-19279 Table 1: Vibrio cholerae Gateway® Clones, Plate 1 (NR-19679) Clone ID Well ORF Locus ID Symbol Product Accession Position Length Number 174071 A02 367 VC2271 ribD riboflavin-specific deaminase NP_231902.1 174346 A03 336 VC1877 lpxK tetraacyldisaccharide 4`-kinase NP_231511.1 174354 A04 342 VC0953 holA DNA polymerase III, delta subunit NP_230600.1 174115 A05 388 VC2085 sucC succinyl-CoA synthase, beta subunit NP_231717.1 174310 A06 506 VC2400 murC UDP-N-acetylmuramate--alanine ligase NP_232030.1 174523 A07 132 VC0644 rbfA ribosome-binding factor A NP_230293.2 174632 A08 322 VC0681 ribF riboflavin kinase-FMN adenylyltransferase NP_230330.1 174930 A09 433 VC0720 phoR histidine protein kinase PhoR NP_230369.1 174953 A10 206 VC1178 conserved hypothetical protein NP_230823.1 174976 A11 213 VC2358 hypothetical protein NP_231988.1 174898 A12 369 VC0154 trmA tRNA (uracil-5-)-methyltransferase NP_229811.1 174059 B01 73 VC2098 hypothetical protein NP_231730.1 174075 B02 82 VC0561 rpsP ribosomal protein S16 NP_230212.1 174087 B03 378 VC1843 cydB-1 cytochrome d ubiquinol oxidase, subunit II NP_231477.1 174099 B04 383 VC1798 eha eha protein NP_231433.1 174294 B05 494 VC0763 GTP-binding protein NP_230412.1 174311 B06 314 VC2183 prsA ribose-phosphate pyrophosphokinase NP_231814.1 174603 B07 108 VC0675 thyA thymidylate synthase NP_230324.1 174474 B08 466 VC1297 asnS asparaginyl-tRNA synthetase NP_230942.2 174933 B09 198 -

Anti-OXCT1 Antibody (ARG58973)
Product datasheet [email protected] ARG58973 Package: 100 μl anti-OXCT1 antibody Store at: -20°C Summary Product Description Rabbit Polyclonal antibody recognizes OXCT1 Tested Reactivity Hu, Ms, Rat Tested Application IHC-P, WB Host Rabbit Clonality Polyclonal Isotype IgG Target Name OXCT1 Antigen Species Human Immunogen Recombinant fusion protein corresponding to aa. 261-520 of Human OXCT1 (NP_000427.1). Conjugation Un-conjugated Alternate Names OXCT; EC 2.8.3.5; Succinyl-CoA:3-ketoacid coenzyme A transferase 1, mitochondrial; SCOT-s; SCOT; Somatic-type succinyl-CoA:3-oxoacid CoA-transferase; 3-oxoacid CoA-transferase 1 Application Instructions Application table Application Dilution IHC-P 1:50 - 1:100 WB 1:500 - 1:2000 Application Note * The dilutions indicate recommended starting dilutions and the optimal dilutions or concentrations should be determined by the scientist. Positive Control Mouse heart Calculated Mw 56 kDa Observed Size 56 kDa Properties Form Liquid Purification Affinity purified. Buffer PBS (pH 7.3), 0.02% Sodium azide and 50% Glycerol. Preservative 0.02% Sodium azide Stabilizer 50% Glycerol Storage instruction For continuous use, store undiluted antibody at 2-8°C for up to a week. For long-term storage, aliquot and store at -20°C. Storage in frost free freezers is not recommended. Avoid repeated freeze/thaw cycles. Suggest spin the vial prior to opening. The antibody solution should be gently mixed before use. www.arigobio.com 1/2 Note For laboratory research only, not for drug, diagnostic or other use. Bioinformation Gene Symbol OXCT1 Gene Full Name 3-oxoacid CoA transferase 1 Background This gene encodes a member of the 3-oxoacid CoA-transferase gene family. -

Supplementary Informations SI2. Supplementary Table 1
Supplementary Informations SI2. Supplementary Table 1. M9, soil, and rhizosphere media composition. LB in Compound Name Exchange Reaction LB in soil LBin M9 rhizosphere H2O EX_cpd00001_e0 -15 -15 -10 O2 EX_cpd00007_e0 -15 -15 -10 Phosphate EX_cpd00009_e0 -15 -15 -10 CO2 EX_cpd00011_e0 -15 -15 0 Ammonia EX_cpd00013_e0 -7.5 -7.5 -10 L-glutamate EX_cpd00023_e0 0 -0.0283302 0 D-glucose EX_cpd00027_e0 -0.61972444 -0.04098397 0 Mn2 EX_cpd00030_e0 -15 -15 -10 Glycine EX_cpd00033_e0 -0.0068175 -0.00693094 0 Zn2 EX_cpd00034_e0 -15 -15 -10 L-alanine EX_cpd00035_e0 -0.02780553 -0.00823049 0 Succinate EX_cpd00036_e0 -0.0056245 -0.12240603 0 L-lysine EX_cpd00039_e0 0 -10 0 L-aspartate EX_cpd00041_e0 0 -0.03205557 0 Sulfate EX_cpd00048_e0 -15 -15 -10 L-arginine EX_cpd00051_e0 -0.0068175 -0.00948672 0 L-serine EX_cpd00054_e0 0 -0.01004986 0 Cu2+ EX_cpd00058_e0 -15 -15 -10 Ca2+ EX_cpd00063_e0 -15 -100 -10 L-ornithine EX_cpd00064_e0 -0.0068175 -0.00831712 0 H+ EX_cpd00067_e0 -15 -15 -10 L-tyrosine EX_cpd00069_e0 -0.0068175 -0.00233919 0 Sucrose EX_cpd00076_e0 0 -0.02049199 0 L-cysteine EX_cpd00084_e0 -0.0068175 0 0 Cl- EX_cpd00099_e0 -15 -15 -10 Glycerol EX_cpd00100_e0 0 0 -10 Biotin EX_cpd00104_e0 -15 -15 0 D-ribose EX_cpd00105_e0 -0.01862144 0 0 L-leucine EX_cpd00107_e0 -0.03596182 -0.00303228 0 D-galactose EX_cpd00108_e0 -0.25290619 -0.18317325 0 L-histidine EX_cpd00119_e0 -0.0068175 -0.00506825 0 L-proline EX_cpd00129_e0 -0.01102953 0 0 L-malate EX_cpd00130_e0 -0.03649016 -0.79413596 0 D-mannose EX_cpd00138_e0 -0.2540567 -0.05436649 0 Co2 EX_cpd00149_e0 -

Succinyl-Coa:3-Ketoacid Coa Transferase Deficiency
Succinyl-CoA:3-ketoacid CoA transferase deficiency Description Succinyl-CoA:3-ketoacid CoA transferase (SCOT) deficiency is an inherited disorder that impairs the body's ability to break down ketones, which are molecules produced in the liver during the breakdown of fats. The signs and symptoms of SCOT deficiency typically appear within the first few years of life. Affected individuals experience episodes of extreme tiredness (lethargy), appetite loss, vomiting, rapid breathing, and, occasionally, seizures. These episodes, which are called ketoacidotic attacks, sometimes lead to coma. About half of affected individuals have a ketoacidotic attack within the first 4 days of life. Affected individuals have no symptoms of the disorder between ketoacidotic attacks. People with SCOT deficiency usually have a permanently elevated level of ketones in their blood (persistent ketosis). If the level of ketones gets too high, which can be brought on by infections, fevers, or periods without food (fasting), a ketoacidotic attack can occur. The frequency of ketoacidotic attacks varies among affected individuals. Frequency The prevalence of SCOT deficiency is unknown. More than 20 cases of this condition have been reported in the scientific literature. Causes Mutations in the OXCT1 gene cause SCOT deficiency. The OXCT1 gene provides instructions for making an enzyme called succinyl-CoA:3-ketoacid CoA transferase ( SCOT). The SCOT enzyme is made in the energy-producing centers of cells ( mitochondria). The enzyme plays a role in the breakdown of ketones, which are an important source of energy during fasting or when energy demands are increased, such as during illness or when exercising. OXCT1 gene mutations result in the production of a SCOT enzyme with little or no function. -

OXCT1 Antibody Cat
OXCT1 Antibody Cat. No.: 30-414 OXCT1 Antibody Antibody used in WB on Hum. Adult Placenta at 1 ug/ml. Specifications HOST SPECIES: Rabbit SPECIES REACTIVITY: Human, Mouse, Rat Antibody produced in rabbits immunized with a synthetic peptide corresponding a region IMMUNOGEN: of human OXCT1. TESTED APPLICATIONS: ELISA, WB OXCT1 antibody can be used for detection of OXCT1 by ELISA at 1:62500. OXCT1 antibody APPLICATIONS: can be used for detection of OXCT1 by western blot at 0.5 μg/mL, and HRP conjugated secondary antibody should be diluted 1:50,000 - 100,000. POSITIVE CONTROL: 1) Cat. No. 1201 - HeLa Cell Lysate PREDICTED MOLECULAR 52 kDa WEIGHT: October 2, 2021 1 https://www.prosci-inc.com/oxct1-antibody-30-414.html Properties PURIFICATION: Antibody is purified by peptide affinity chromatography method. CLONALITY: Polyclonal CONJUGATE: Unconjugated PHYSICAL STATE: Liquid Purified antibody supplied in 1x PBS buffer with 0.09% (w/v) sodium azide and 2% BUFFER: sucrose. CONCENTRATION: batch dependent For short periods of storage (days) store at 4˚C. For longer periods of storage, store STORAGE CONDITIONS: OXCT1 antibody at -20˚C. As with any antibody avoid repeat freeze-thaw cycles. Additional Info OFFICIAL SYMBOL: OXCT1 ALTERNATE NAMES: OXCT1, OXCT, SCOT ACCESSION NO.: NP_000427 PROTEIN GI NO.: 4557817 GENE ID: 5019 USER NOTE: Optimal dilutions for each application to be determined by the researcher. Background and References OXCT1 is a member of the 3-oxoacid CoA-transferase gene family. It is a homodimeric mitochondrial matrix enzyme that plays a central role in extrahepatic ketone body catabolism by catalyzing the reversible transfer of coenzyme A from succinyl-CoA to acetoacetate.This gene encodes a member of the 3-oxoacid CoA-transferase gene family. -

OXCT1 (NM 000436) Human Recombinant Protein – TP303764
OriGene Technologies, Inc. 9620 Medical Center Drive, Ste 200 Rockville, MD 20850, US Phone: +1-888-267-4436 [email protected] EU: [email protected] CN: [email protected] Product datasheet for TP303764 OXCT1 (NM_000436) Human Recombinant Protein Product data: Product Type: Recombinant Proteins Description: Recombinant protein of human 3-oxoacid CoA transferase 1 (OXCT1), nuclear gene encoding mitochondrial protein Species: Human Expression Host: HEK293T Tag: C-Myc/DDK Predicted MW: 52 kDa Concentration: >50 ug/mL as determined by microplate BCA method Purity: > 80% as determined by SDS-PAGE and Coomassie blue staining Buffer: 25 mM Tris.HCl, pH 7.3, 100 mM glycine, 10% glycerol Preparation: Recombinant protein was captured through anti-DDK affinity column followed by conventional chromatography steps. Storage: Store at -80°C. Stability: Stable for 12 months from the date of receipt of the product under proper storage and handling conditions. Avoid repeated freeze-thaw cycles. RefSeq: NP_000427 Locus ID: 5019 UniProt ID: P55809, A0A024R040 RefSeq Size: 3572 Cytogenetics: 5p13.1 RefSeq ORF: 1560 Synonyms: OXCT; SCOT Summary: This gene encodes a member of the 3-oxoacid CoA-transferase gene family. The encoded protein is a homodimeric mitochondrial matrix enzyme that plays a central role in extrahepatic ketone body catabolism by catalyzing the reversible transfer of coenzyme A from succinyl-CoA to acetoacetate. Mutations in this gene are associated with succinyl CoA:3- oxoacid CoA transferase deficiency. [provided by RefSeq, Jul 2008] This product is to be used for laboratory only. Not for diagnostic or therapeutic use. View online » ©2021 OriGene Technologies, Inc., 9620 Medical Center Drive, Ste 200, Rockville, MD 20850, US 1 / 2 OXCT1 (NM_000436) Human Recombinant Protein – TP303764 Protein Pathways: Butanoate metabolism, Synthesis and degradation of ketone bodies, Valine, leucine and isoleucine degradation Product images: Coomassie blue staining of purified OXCT1 protein (Cat# TP303764).