The Hunt for Natural Skin Whitening Agents
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Thesis of Potentially Sweet Dihydrochalcone Glycosides
University of Bath PHD The synthesis of potentially sweet dihydrochalcone glycosides. Noble, Christopher Michael Award date: 1974 Awarding institution: University of Bath Link to publication Alternative formats If you require this document in an alternative format, please contact: [email protected] General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal ? Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Download date: 05. Oct. 2021 THE SYNTHESIS OF POTBTTIALLY SWEET DIHYDROCHALCOITB GLYCOSIDES submitted by CHRISTOPHER MICHAEL NOBLE for the degree of Doctor of Philosophy of the University of Bath. 1974 COPYRIGHT Attention is drawn to the fact that copyright of this thesis rests with its author.This copy of the the sis has been supplied on condition that anyone who con sults it is understood to recognise that its copyright rests with its author and that no quotation from the thesis and no information derived from it may be pub lished without the prior written consent of the author. -

Treatment Protocol Copyright © 2018 Kostoff Et Al
Prevention and reversal of Alzheimer's disease: treatment protocol Copyright © 2018 Kostoff et al PREVENTION AND REVERSAL OF ALZHEIMER'S DISEASE: TREATMENT PROTOCOL by Ronald N. Kostoffa, Alan L. Porterb, Henry. A. Buchtelc (a) Research Affiliate, School of Public Policy, Georgia Institute of Technology, USA (b) Professor Emeritus, School of Public Policy, Georgia Institute of Technology, USA (c) Associate Professor, Department of Psychiatry, University of Michigan, USA KEYWORDS Alzheimer's Disease; Dementia; Text Mining; Literature-Based Discovery; Information Technology; Treatments Prevention and reversal of Alzheimer's disease: treatment protocol Copyright © 2018 Kostoff et al CITATION TO MONOGRAPH Kostoff RN, Porter AL, Buchtel HA. Prevention and reversal of Alzheimer's disease: treatment protocol. Georgia Institute of Technology. 2018. PDF. https://smartech.gatech.edu/handle/1853/59311 COPYRIGHT AND CREATIVE COMMONS LICENSE COPYRIGHT Copyright © 2018 by Ronald N. Kostoff, Alan L. Porter, Henry A. Buchtel Printed in the United States of America; First Printing, 2018 CREATIVE COMMONS LICENSE This work can be copied and redistributed in any medium or format provided that credit is given to the original author. For more details on the CC BY license, see: http://creativecommons.org/licenses/by/4.0/ This work is licensed under a Creative Commons Attribution 4.0 International License<http://creativecommons.org/licenses/by/4.0/>. DISCLAIMERS The views in this monograph are solely those of the authors, and do not represent the views of the Georgia Institute of Technology or the University of Michigan. This monograph is not intended as a substitute for the medical advice of physicians. The reader should regularly consult a physician in matters relating to his/her health and particularly with respect to any symptoms that may require diagnosis or medical attention. -

Chromanone-A Prerogative Therapeutic Scaffold: an Overview
Arabian Journal for Science and Engineering https://doi.org/10.1007/s13369-021-05858-3 REVIEW-CHEMISTRY Chromanone‑A Prerogative Therapeutic Scafold: An Overview Sonia Kamboj1,2 · Randhir Singh1 Received: 28 September 2020 / Accepted: 9 June 2021 © King Fahd University of Petroleum & Minerals 2021 Abstract Chromanone or Chroman-4-one is the most important and interesting heterobicyclic compound and acts as a building block in medicinal chemistry for isolation, designing and synthesis of novel lead compounds. Structurally, absence of a double bond in chromanone between C-2 and C-3 shows a minor diference from chromone but exhibits signifcant variations in biological activities. In the present review, various studies published on synthesis, pharmacological evaluation on chroman- 4-one analogues are addressed to signify the importance of chromanone as a versatile scafold exhibiting a wide range of pharmacological activities. But, due to poor yield in the case of chemical synthesis and expensive isolation procedure from natural compounds, more studies are required to provide the most efective and cost-efective methods to synthesize novel chromanone analogs to give leads to chemistry community. Considering the versatility of chromanone, this review is designed to impart comprehensive, critical and authoritative information about chromanone template in drug designing and development. Keywords Chroman-4-one · Chromone · Pharmacological activity · Synthesis · Analogues 1 Introduction dihydropyran (ring B) which relates to chromane, chromene, chromone and chromenone, but the absence of C2-C3 dou- Chroman-4-one is one of the most important heterobicyclic ble bond of chroman-4-one skeleton makes a minor difer- moieties existing in natural compounds as polyphenols and ence (Table 1) from chromone and associated with diverse as synthetic compounds like Taxifolin, also known as chro- biological activities [1]. -

Flavonoid Glucodiversification with Engineered Sucrose-Active Enzymes Yannick Malbert
Flavonoid glucodiversification with engineered sucrose-active enzymes Yannick Malbert To cite this version: Yannick Malbert. Flavonoid glucodiversification with engineered sucrose-active enzymes. Biotechnol- ogy. INSA de Toulouse, 2014. English. NNT : 2014ISAT0038. tel-01219406 HAL Id: tel-01219406 https://tel.archives-ouvertes.fr/tel-01219406 Submitted on 22 Oct 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Last name: MALBERT First name: Yannick Title: Flavonoid glucodiversification with engineered sucrose-active enzymes Speciality: Ecological, Veterinary, Agronomic Sciences and Bioengineering, Field: Enzymatic and microbial engineering. Year: 2014 Number of pages: 257 Flavonoid glycosides are natural plant secondary metabolites exhibiting many physicochemical and biological properties. Glycosylation usually improves flavonoid solubility but access to flavonoid glycosides is limited by their low production levels in plants. In this thesis work, the focus was placed on the development of new glucodiversification routes of natural flavonoids by taking advantage of protein engineering. Two biochemically and structurally characterized recombinant transglucosylases, the amylosucrase from Neisseria polysaccharea and the α-(1→2) branching sucrase, a truncated form of the dextransucrase from L. Mesenteroides NRRL B-1299, were selected to attempt glucosylation of different flavonoids, synthesize new α-glucoside derivatives with original patterns of glucosylation and hopefully improved their water-solubility. -

Producing Cultured Cells of Sophora Flavescens
Plant Biotechnology 21(5), 355–359 (2004) Original Paper Metabolism of administered (2RS)-naringenin in flavonoid- producing cultured cells of Sophora flavescens Hirobumi Yamamoto*, Hiroki Kuribayashi, Yasuharu Seshima, Ping Zhao1, Isao Kouno1, Goro Taguchi2, Koichiro Shimomura Plant Regulation Research Center, Faculty of Life Sciences, Toyo University, 1-1-1 Izumino, Itakura-machi, Oura, Gunma 374-0193, Japan 1Medicinal Plant Garden, Course of Pharmaceutical Sciences, Graduate School of Biomedical Sciences, Nagasaki University, 1-14 Bunkyo-machi, Nagasaki 852-8521, Japan 2Division of Gene Research, Research Center for Human and Environmental Sciences, Shinshu University, 3-15-1 Tokida, Nagano 386-8567, Japan *E-mail: [email protected] Tel: +81-276-82-9206 Fax: +81-276-82-9206 Received August 30, 2004; accepted October 4, 2004 (Edited by K. Yazaki) Abstract Cultured cells of Sophora flavescens produce (2S)-naringenin-derived prenylated flavanone sophoraflavanone G and liquiritigenin-derived trifolirhizin 6Ј-O-malonate. The regulation of flavonoid biosynthesis was examined by analyzing the metabolites produced in the cultured cells fed (2RS)-naringenin. The amount of sophoraflavanone G in cells fed 0.1 or 0.3 mM (2RS)-naringenin was two-fold that in control cells, although the conversion ratio was only 5 to 10% of the administered (2S)-naringenin. On the other hand, (2R)-naringenin, which does not occur naturally, was efficiently converted into its 4Ј,7-di-O-b-D-glucoside. (2S)-Naringenin prenylation activity was higher at the logarithmic growth stage. The cells fed (2RS)-naringenin at a lower concentration (below 0.1 mM), accumulated sophoraflavanone G as the main prenylated flavanone. -

In Chemistry, Glycosides Are Certain Molecules in Which a Sugar Part Is
GLYCOSIDES Glycosides may be defined as the organic compounds from plants or animal sources, which on enzymatic or acid hydrolysis give one or more sugar moieties along with non- sugar moiety. Glycosides play numerous important roles in living organisms. Many plants store important chemicals in the form of inactive glycosides; if these chemicals are needed, the glycosides are brought in contact with water and an enzyme, and the sugar part is broken off, making the chemical available for use. Many such plant glycosides are used as medications. In animals (including humans), poisons are often bound to sugar molecules in order to remove them from the body. Formally, a glycoside is any molecule in which a sugar group is bonded through its carbon atom to another group via an O-glycosidic bond or an S-glycosidic bond; glycosides involving the latter are also called thioglycosides. The sugar group is then known as the glycone and the non-sugar group as the aglycone or genin part of the glycoside. The glycone can consist of a single sugar group (monosaccharide) or several sugar groups (oligosaccharide). Classification Classification based on linkages Based on the linkage of sugar moiety to aglycone part 1. O-Glycoside:-Here the sugar is combined with alcoholic or phenolic hydroxyl function of aglycone.eg:-digitalis. 2. N-glycosides:-Here nitrogen of amino group is condensed with a sugar ,eg- Nucleoside 3. S-glycoside:-Here sugar is combined with sulphur of aglycone,eg- isothiocyanate glycosides. 4. C-glycosides:-By condensation of a sugar with a cabon atom, eg-Cascaroside, aloin. Glycosides can be classified by the glycone, by the type of glycosidic bond, and by the aglycone. -

Nine Traditional Chinese Herbal Formulas for the Treatment of Depression: an Ethnopharmacology, Phytochemistry, and Pharmacology Review
Journal name: Neuropsychiatric Disease and Treatment Article Designation: Review Year: 2016 Volume: 12 Neuropsychiatric Disease and Treatment Dovepress Running head verso: Feng et al Running head recto: Chinese herbal formulas as antidepressants open access to scientific and medical research DOI: http://dx.doi.org/10.2147/NDT.S114560 Open Access Full Text Article REVIEW Nine traditional Chinese herbal formulas for the treatment of depression: an ethnopharmacology, phytochemistry, and pharmacology review Dan-dan Feng Abstract: Depression is a major mental disorder, and is currently recognized as the Tao Tang second-leading cause of disability worldwide. However, the therapeutic effect of antidepressants Xiang-ping Lin remains unsatisfactory. For centuries, Chinese herbal formulas (CHFs) have been widely used in Zhao-yu Yang the treatment of depression, achieving better therapeutic effects than placebo and having fewer Shu Yang side effects than conventional antidepressants. Here, we review the ethnopharmacology, phy- Zi-an Xia tochemistry, and pharmacology studies of nine common CHFs: “banxia houpo” decoction, “chaihu shugansan”, “ganmaidazao” decoction, “kaixinsan”, “shuganjieyu” capsules, “sinisan”, Yun Wang “wuling” capsules, “xiaoyaosan”, and “yueju”. Eight clinical trials and seven meta-analyses have Piao Zheng supported the theory that CHFs are effective treatments for depression, decreasing Hamilton Yang Wang Depression Scale scores and showing few adverse effects. Evidence from 75 preclinical studies Chun-hu Zhang has also elucidated the multitarget and multipathway mechanisms underlying the antidepres- Laboratory of Ethnopharmacology, sant effect of the nine CHFs. Decoctions, capsules, and pills all showed antidepressant effects, Institute of Integrated Traditional ranked in descending order of efficacy. According to traditional Chinese medicine theory, these Chinese and Western Medicine, Xiangya Hospital, Central South CHFs have flexible compatibility and mainly act by soothing the liver and relieving depression. -

View PDF Version
RSC Advances View Article Online PAPER View Journal | View Issue Comparative intestinal bacteria-associated pharmacokinetics of 16 components of Shengjiang Cite this: RSC Adv.,2017,7,43621 Xiexin decoction between normal rats and rats with irinotecan hydrochloride (CPT-11)-induced gastrointestinal toxicity in vitro using salting-out sample preparation and LC-MS/MS† Huanyu Guan, ab Xiaoming Wang,a Shiping Wang,b Yang He,a Jiajing Yue,a Shanggao Liao,b Yuanda Huanga and Yue Shi*a Shengjiang Xiexin decoction (SXD) exerts protective effects against gastrointestinal injury induced by irinotecan hydrochloride (CPT-11). The intestinal bacteria-associated in vitro pharmacokinetics of 16 components of SXD in normal rats and those with CPT-11-induced gastrointestinal toxicity were Creative Commons Attribution 3.0 Unported Licence. compared in this study. A sensitive and reproducible ultra-high-performance liquid chromatography coupled to tandem mass spectrometry (UHPLC-MS/MS) method was developed for the quantification of 16 components of SXD in a rat intestinal bacteria incubation system, using naringin, naringenin and tetrahydropalmatine as internal standards (ISs). The samples were prepared via salting-out assisted liquid–liquid extraction (SALLE) with NaCl to reduce matrix effects. Chromatographic separation was performed on a sub-2 mm analytical column with acetonitrile and 0.1% aqueous formic acid as mobile phase. All of the analyzed components and ISs were detected via multiple reaction monitoring (MRM) scanning with electrospray ionization. The proposed method was successfully applied for the in vitro This article is licensed under a pharmacokinetic analysis of the multiple components of a complex mixture consisting of a traditional Chinese medicine (TCM) and an intestinal bacterial incubation system. -

Cushnie TPT, Lamb AJ. Antimicrobial Activity of Flavonoids. International Journal of Antimicrobial Agents, 2005. 26(5):343-356
Cushnie TPT, Lamb AJ. Antimicrobial activity of flavonoids. International Journal of Antimicrobial Agents, 2005. 26(5):343-356. PMID: 16323269 DOI: 10.1016/j.ijantimicag.2005.09.002 The journal article above is freely available from the publishers at: http://www.idpublications.com/journals/PDFs/IJAA/ANTAGE_MostCited_1.pdf and also... http://www.ijaaonline.com/article/S0924-8579(05)00255-4/fulltext Errata for the article (typesetting errors by Elsevier Ireland) are freely available from the publishers at: http://www.ijaaonline.com/article/S0924-8579(05)00352-3/fulltext and also... http://www.sciencedirect.com/science/article/pii/S0924857905003523 International Journal of Antimicrobial Agents 26 (2005) 343–356 Review Antimicrobial activity of flavonoids T.P. Tim Cushnie, Andrew J. Lamb ∗ School of Pharmacy, The Robert Gordon University, Schoolhill, Aberdeen AB10 1FR, UK Abstract Flavonoids are ubiquitous in photosynthesising cells and are commonly found in fruit, vegetables, nuts, seeds, stems, flowers, tea, wine, propolis and honey. For centuries, preparations containing these compounds as the principal physiologically active constituents have been used to treat human diseases. Increasingly, this class of natural products is becoming the subject of anti-infective research, and many groups have isolated and identified the structures of flavonoids possessing antifungal, antiviral and antibacterial activity. Moreover, several groups have demonstrated synergy between active flavonoids as well as between flavonoids and existing chemotherapeutics. Reports of activity in the field of antibacterial flavonoid research are widely conflicting, probably owing to inter- and intra-assay variation in susceptibility testing. However, several high-quality investigations have examined the relationship between flavonoid structure and antibacterial activity and these are in close agreement. -

An Investigation on the Anti-Tumor Activities of Sophoraflavanone G on Human Myeloid Leukemia Cells
An Investigation on the Anti-tumor Activities of Sophoraflavanone G on Human Myeloid Leukemia Cells LIU, Xiaozhuo A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Philosophy in Biochemistry • The Chinese University of Hong Kong September 2008 The Chinese University of Hong Kong holds the copyright of this thesis. Any person(s) intending to use a part or whole of the materials in the thesis in a proposed publication must seek copyright release from the Dean of the Graduate School. 統系餘t圖 |( 2 0 1 雇)l) ^^ UNIVERSITY /M ^^S^UBRARY SYSTEMX^ Declaration I declare that the assignment here submitted is original except for source material explicitly acknowledged. I also acknowledge that I am aware of University policy and regulations on honesty in academic work, and of the disciplinary guidelines and procedures applicable to breaches of such policy and regulations, as contained in the website http://www.cuhk.edu.hk/policy/academichonesty/ 16/09/2008 Signature Date LIU Xiaozhuo 06148340 Name Student ID Thesis of M.Phil, in Biochemistry Title: An Investigation on the Anti-tumor Activities of Sophoraflavanone G on Human Myeloid Leukemia Cells Course code Course title Thesis/ Assessment Committee Professor S. K. Kong (Chair) Professor K. N. Leung (Thesis Supervisor) Professor K. P. Fung (Thesis Supervisor) Professor C. K. Wong (Committee Member) Professor Ricky N. S. Wong (External Examiner) Abstract Abstract During the last one and a half centuries, our basic understanding of leukemia has expanded greatly, from the first description as "white blood" in the year of 1845 to the recognition of leukemia as disorders of hematopoietic cell differentiation and apoptosis. -
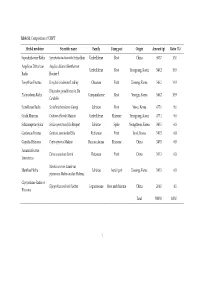
Ratio (%) Saposhnikoviae Ra
Table S1. Composition of CSBPT Herbal medicine Scientific name Family Using part Origin Amount (g) Ratio (%) Saposhnikoviae Radix Saposhnikovia divaricate Schischkin Umbelliferae Root China 500.7 10.0 Angelicae Dahuricae Angelica dahurica Bentham et Umbelliferae Root Yeongyang, Korea 546.2 10.9 Radix Hooker F. Forsythiae Fructus Forsythia viridissima Lindley Oleaceae Fruit Uiseong, Korea 546.2 10.9 Platycodon grandiflorum A. De Platycodonis Radix Campanulaceae Root Yeongju, Korea 546.2 10.9 Candolle Scutellariae Radix Scutellaria baicalensis Georgi Labiatae Root Yeosu, Korea 477.1 9.6 Cnidii Rhizoma Cnidium officinale Makino Umbelliferae Rhizome Yeongyang, Korea 477.1 9.6 Schizonepetae Spica Schizonepeta tenuifolia Briquet Labiatae Spike Yeongcheon, Korea 340.5 6.8 Gardeniae Fructus Gardenia jasminoides Ellis Rubiaceae Fruit Imsil, Korea 340.5 6.8 Coptidis Rhizoma Coptis japonica Makino Ranunculaceae Rhizome China 340.5 6.8 Aurantii Fructus Citrus aurantium Linné Rutaceae Fruit China 340.5 6.8 Immaturus Mentha arvensis Linné var. Menthae Herba Labiatae Aerial part Uiseong, Korea 340.5 6.8 piperascens Malinvaud ex Holmes Glycyrrhizae Radix et Glycyrrhiza uralensis Fischer Leguminosae Root and rhizome China 204.0 4.1 Rhizoma Total 5000.0 100.0 1 Table 2. Chromatographic conditions for simultaneous quantification of compounds 1–18 in CSBPT. Chromatographic parameter Column SunFire C18 analytical column (250 × 4.6 mm, 5 μm) Detector PDA (235, 250, 280, 310, and 345 nm) Flow rate (mL/min) 1.0 Injection volume (μL) 10.0 Column temperature (°C) 40.0 A: 0.1% (v/v) aqueous formic acid Mobile phase B: 0.1% (v/v) formic acid in acetonitrile Time (min) A (%) B (%) 0 95 5 40 40 60 Gradient elution 50 5 95 55 5 95 60 95 5 70 95 5 2 Table S3. -

Simultaneous Determination of Nine Bioactive
Accepted Manuscript Title: Simultaneous determination of nine bioactive compounds in Yijin-tang via high-performance liquid chromatography and liquid chromatography-electrospray ionization-mass spectrometry Author: Hye Jin Yang Nam-Hui Yim Kwang Jin Lee Min Jung Gu Bohyoung Lee Youn-Hwan Hwang Jin Yeul Ma PII: S2213-4220(16)30030-0 DOI: http://dx.doi.org/doi:10.1016/j.imr.2016.04.005 Reference: IMR 200 To appear in: Received date: 14-10-2015 Revised date: 30-3-2016 Accepted date: 7-4-2016 Please cite this article as: Hye Jin YangNam-Hui YimKwang Jin LeeMin Jung GuBohyoung LeeYoun-Hwan HwangJin Yeul Ma Simultaneous determination of nine bioactive compounds in Yijin-tang via high-performance liquid chromatography and liquid chromatography-electrospray ionization-mass spectrometry (2016), http://dx.doi.org/10.1016/j.imr.2016.04.005 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. 1 Original Article 2 3 Simultaneous determination of nine bioactive compounds in Yijin-tang via high- 4 performance liquid chromatography and liquid chromatography-electrospray 5 ionization-mass spectrometry 6 7 Hye Jin Yang, Nam-Hui Yim, Kwang Jin Lee, Min Jung Gu, Bohyoung Lee, Youn-Hwan * * 8 Hwang and Jin Yeul Ma 9 10 KM Application Center, Korea Institute of Oriental Medicine, 11 70 Cheomdanro, Dong-gu, Daegu 701-300, Republic of Korea 12 13 Running title: Simultaneous determination of Yijin-tang via HPLC-DAD and LC/MS 14 * 15 Address for co-corresponding author 16 Youn-Hwan Hwang, Ph.D.