Innledning, Finale
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hvordan Er Den Naturlige Rekrutteringa Til Røye I Øvre Og Nedre Roasten?
Campus Evenstad Skog og utmarksfag Andreas Frøsland Hvordan er den naturlige rekrutteringa til røye i Øvre og Nedre Roasten? Bacheloroppgave i utmarksforvaltning 2010 Utlånsklausul: Nei Ja. Antall år Høgskolen i Hedmark Høgskoleadministrasjonen Postadresse: 2418 Elverum Besøksadresse: Lærerskolealléen 1 Telefon +47 62 43 00 00 Telefaks +47 62 43 00 01 E-post [email protected] Org. nr 974 251 760 www.hihm.no Forord Dette er min avsluttende oppgave i min bachelorutdannelse her på Evenstad. Målet med oppgaven er å se på den naturlige rekrutteringa til Røya i øvre og nedre Roasten i Femundsmarka nasjonalpark. Arbeidet er blitt utført på oppdrag fra Engerdal fjellstyre og Statsskog på Røros. Bakgrunnen for oppgaven er å se om røyebestanden er balansert i forhold til det sportsfiske og garnfiske som blir utført i området. Det har vært bekymringer om røyebestanden grunnet høyt fiskepress. Jeg ville også se på om utsetting av røye i Roastensjøene hadde noen effekt på den totale fiskebestanden. Oppgaven har vært lang og tidkrevende. Det har vært fysisk krevende med mye bæring og korte netter. Jeg vil takke min veileder Kjell Langdal for oppfølgingen og veiledningen under prosjektet. Jeg vil takke Høgskolen i Hedmark avd. Evenstad for utlåning av utstyr og Evenstad settefiskanlegg for utlåning av ruser. Jeg vil også takke Engerdal fjellstyre og spesielt Torfinn Slettevold for hans hjelp og tålmodighet under vår kartlegging av dybdeforhold i Roastensjøene. I tillegg vil jeg takke Statskog på Røros for utlån av båt og husly under prøvefisket og til slutt vil jeg rette en spesiell takk til Geir Wagnild for hjelp og veiledning under feltarbeidet. -

Statskogs Åpne Buer I Femundsmarka
Statskogs åpne buer i Femundsmarka FERAGEN Langtjønna landskapsvernområde Fjølburøsta Ljøsnåvollen 648 654 Langtjønnbua Femundsåsen Furubakken Harrbekkbua 783 Muggsjølia Litjrennbua Langen Sellesdalsbua Lorthølbua Muggsjøbua Grunnhåbua Grådalsbua S Langmyrbua V E Flensmarka Gubbtjønnbua naturreservat Fautbua R 662 780 I G FEMUNDEN E 1292 759 Flenskampan FEMUNDSMARKA NASJONALPARK Røosbua Røvollen Revbua Roastkoia Møllerbua Kløfthåbua Fløtarbua ved Roasten 783 1416 788 Store Svuku 662 Femundslia landskapsvernområde Svukuriset H2y8tter, jakt, fiske og friluftsliv Tegnforklaring 888 - Oversikt over Statskogs tilbud: statskog.no Åpen bu - Jakt- og fiskekort samt hytteutleie: inatur.no nasjonalpark - Flotte turforslag og kart: godtur.no landskapsvernområde - Info/kjøpshjelp på friluftstorget: tlf 07800 (tast 1) naturreservat 0 1,25 2,5 5 Kilometer Kartografi: JC Torvik 221 1460 VELKOMMEN! De åpne buene i Femundsmarka er kulturminner. De ble i sin tid bygd og brukt som husvære for skogsarbeidere og fløtere. Noen av dem er også gamle seterbuer, andre ble satt opp som reine jakt- og fiskebuer. I dag eies de fleste av de åpne buene i Femundsmarka av Statskog. Statskog har som mål å bevare buene i sin opprinnelige form og i sin opprinnelige funksjon som enkle overnattingsplasser for folk som ferdes i marka. Buene holdes derfor åpne for allmennheten. Hvis du ønsker å fordype deg ytterligere i Femundsmarkas historie, finner du mye interessant stoff bl.a. i følgende publikasjoner: y Buer i Femundsmarka. Røros Museums- og historielag. Røros 2002. y Stor-Hans. Eneboeren i Femundsmarka. Røros Museums- og historielag. Røros 1996. y Fløtningshistorie i Femundsmarka. Med fløterne og tømmerstokkene fra Rogen til Gjøsvika. Røros kommune. Røros 1997. KONTAKT: Statskog, tlf. sentralbord: 74 21 30 00 Statens naturoppsyn (SNO) v/ Tom Johansen tlf: 404 11 426 Ulvelav FAUTBUA Denne bua ble satt opp i 1918 av Jens Jensen Langen (kalt ”litj-fauten”), som var reinoppsyn. -

Opprettelse Av Dåapma Nasjonalpark
Fylkesmannen i Sør-Trøndelag Opprettelse av Dåapma nasjonalpark Revidering av forskrifter for Tekssjøen og Finnvolldalen - Esplingdalen naturreservater Høringsforslag Rapport Miljøvernavd. nr. 2012/5 Fylkesmannen Fylkesmannen i Nord-Trøndelag i Sør-Trøndelag Miljøvernavdelingene R A P P O R T 2012/5 TITTEL DATO: Opprettelse av Dåapma nasjonalpark 2012-12-06 Revidering av forskrifter for Tekssjøen og Finnvolldalen – Esplingdalen naturreservater Høringsforslag FORFATTER ANT. SIDER: Senioringeniør Eldar Ryan 121 Seniorrådgiver Jan-Erik Andersen AVDELING/ENHET ANSV. SIGN: Fylkesmannen i Nord-Trøndelag Miljøverndirektør Svein Karlsen Fylkesmannen i Sør-Trøndelag Miljøverndirektør Stein-Arne Andreassen Miljøvernavdelingene EKSTRAKT Rapporten er høringsforslag for opprettelse av Dåapma nasjonalpark og revidering av forskriftene for Tekssjøen naturreservat og Finnvolldalen-Esplingdalen naturreservat. Området ligger i Åfjord og Osen kommune i Sør-Trøndelag, og Namdalseid kommune i Nord-Trøndelag. Området er nasjonalt verneverdig, og utfyller mangler i den norske nasjonalparkplanen som et lavalpint, kystnært, oseanisk fjellområde. Området er relativt urørt og utgjør det største inngrepsfrie naturområdet på Fosen. Området har store skogbiologiske verdier, viktige ornitologiske verdier, og utgjør et viktig til svært viktig friluftsområde. Det er mange samiske kulturminner i området. De viktigste brukerinteresser er reindrift, beitebruk og friluftsliv. S T I K K O R D Nasjonalpark Naturreservat Dåapma Tekssjøen Finnvolldalen – Esplingdalen 1 Forord Bakgrunnen for høringsforslaget er et initiativ fra de aktuelle kommuner til Miljøverndepartementet i september 2009, hvor det ble fremmet ønske om utredning av mulig nasjonalpark på Nord-Fosen. Fra initiativet kom, har det nå tatt drøyt 2 ½ år å innhente materiale, bearbeide saken og utforme et verneforslag. Saken er underveis bearbeidet og drøftet både i et Arbeidsutvalg med representanter fra samene, kommunene og fylkeskommunene og i en Referansegruppe med representanter fra sentrale parter i prosessen. -

Kraf Tutbygging-Naturvern
RAPPORT fra kontaktutvalget Kraf tutbygging-naturvern om vassdrag som bar vernes mot krafiutbygging Trykt i Reklaetrykk A.s 13ergen 1971 á Til Det k angel ige' departemen t for industri og håndverk Det kongelige kommunal- og arbeidsdepartement Hovedstyret for Norges vassdrags- og elekrrisitetscesen Kontaktutvalget mtbygging-natumern, sosl hzw hatt I oppdrag B utarbeide en Ikte over vasshg som b0r unntas fra mtbygging, har avgitt sin rap- port 30. desember 1970. Som bilag tii rapporten felger : l) Kart over nedbrfeitene til de Passdrag som ~~ unntatt fra -t- bygging. 2) Kamt over vassdrag med nedbarfelt stcarre enn 100 W, sum er uber~rteeller ubetydelig bemrte av kraftutbygging. Som trykte vedlegg felger : 1. uFmel8pig im&dling om natumitenskapelige Mer- ved vassdragsrefl- leringer, Universitetet i Oslos. (Utarbeidet av cand.real. Jan Abrahamsen.) 2. a!i?iigang på og etterspørsel etter elakkhk kraft+. Notat fra fin8n5rBd Eivind Erichsen, Som utrykte vediegg f0lger innkomne utWW b: 1) Fykkemwnnene 2) Dixktowitet for jakt, viitstell og ferskvandiske 3) Den Node Turi&ore.ning og andm turktfo&ger m.v. 4) Uttalekm fra lukalforeninger formidlet gjennom Norgee Jeger- og FWzer- forbund 5) Luktalforeninger av Norges Naturvernforbund 6) Pmfastor Axel SemPee (Av praktiske hensyn er tidlige trykket materiale, fotografier og kartbilag ilsike tatt d i det utrykte vedlegget.) Ham P. Sperstad Vassd~~gdrektør Dosent, dr. Edre Elgmrk Fifiawdd Eivind Erichen Kontaktutvdget for vassdragareg. Stateas Frihdtsrgd Universitetet i Oslo Udmdirektm Gunnar Germeten Professor, dr. Olav Gj~revoZt Kommunaldepa&ment et Stateras Naturvernråd La&luzpsarkitekt Knut Ove Hilkstd Ptsgajef Ernst H. G. Wessel m NVE á INNHOLD Side Side INNLEDNING . l .4. -

Stedsnavn I Nord-Østerdal
Stedsnavn i Nord-Østerdal JON OLAV RYEN Stedsnavn i Nord-Østerdal med Rondane, Rørosfjella og Femundstraktene KOLOFON FORLAG © Jon Olav Ryen Stedsnavn i Nord-Østerdal med Rondane, Rørosfjella og Femundstraktene Kolofon Forlag AS 2015 Forside- og baksidefoto: Rondane, med Midtronden og Digerronden (foto: Geir Olav Slåen). Prosjektet produseres på oppdrag fra forfatter/oppdragsgiver/utgiver Jon Olav Ryen Alle rettigheter/ansvar for prosjektets innhold tillegges Jon Olav Ryen Henvendelser utover bestilling av produktet bes rettet til Jon Olav Ryen ISBN 978-82-300-1287-1 Produksjon: Kolofon Forlag AS, 2015 Boken kan kjøpes i bokhandelen eller www.kolofon.com Det må ikke kopieres fra denne boken i strid med Åndsverkloven eller avtaler om kopiering inngått med KOPINOR, interesseorgan for rettighetshavere til åndsverk. Kopiering i strid med lov eller avtale medfører erstatningsansvar og inndraging og kan straffes med bøter eller fengsel. Innhold Forord ..................................................... 7 Innledning ............................................. 8 Bakgrunn, formål og metode ................. 8 Utvalg av stedsnavn ............................... 10 Skrivemåte og oppsett ............................ 12 Henvisninger, bruk av kilder ................ 13 Lydskrift .................................................. 14 Forkortelser, forklaringer ...................... 15 Stedsnavn .............................................. 17 Sørsamiske terrengord ....................... 483 Kilder .................................................... -
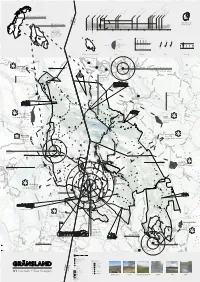
01 Context + Site Analysis
ing techniques d ish side orks orks ing by the Sami RIEN d os Copper W eindeer her os Copper W ing to come ines due to övelsjön opens oject e start ee l icated r ion Pr ease in farming on the Swed Rising tr 2009-2011 Gränslandet End of the last Ice Age Domest Mountain farms 1644 Start of Rør Falcon CatchingCarl Linneaus visits Gränslandet Incr Tourists ar 1926 DNT1937 Svukuriset STF GrChange opens in Sami lifestyle:1977 modern Closing housing of Røimte rand changeher cl Informat 01 Norway & Sweden 835 500 km2 Mountain farms Cattle herding Rogen Naturreservat is Logging and log driving Lumberjack cabins NORWAY home to Swedens only 02 Gränslandet SWEDEN tribe of wild muskox. 2 Ramundberget44 km radius 2000 km “circumference right” 9 areas of protected AURSUNDEN nature situated on the ago 1600 1700 1800 1900 2000 southern tip of the years Scandinavian Mountain 000 8000 years ago range, crossing over the 10 border of Norway and Brekken Messlingen Sweden. Røros 29.6 °C 30.1 °C 28.7 °C 29.3 °C SPRING mid April - mid June Tännäs Bruksvallarna Yearly average 1.6 °C 1.36 m ÖVER-SÄRVSJÖN Hitterdal WINTER SUMMER 1.0 m November - mid April mid June - September 0.5 m 1. 2.8 m/s 2. 8 m/s 3. 38.1 m/s 0.2 m -28.8 °C -31.4 °C SW 197° Fjällnäs -38 °C -37.1 °C l Drevsjø AUTUMN l os r Idre Fjäll September - November evsjø Rø ännäs r e Fjä r T D Id Weather stations for data collection Seasons Temperature Wind Snow depth Langtjønna FERAGEN Tänndalen LANDSKAPSVERNOMRÅDE Storvigeln Storvigeln since 2003 1561 MASL Fjällnäs, Tänndalen, Funäsdalen, Tännäs Highest mountain top Funäsdalen protected area OLD SAMI HUT Villages north of Gränslandet. -
Femundsmarka
FEMUNDSMARKA With a canoe and a fishing rod to Norway’s morning land 2° 3° Femundsmarka National Park Femundsmarka National Park The gift from the ice With twisted pines and secluded lakes scattered amongst a sea of boulders, the Femundsmarka National Park beckons you to experience it. Ten thousand years ago, the glaciers retreated leaving behind a desolate, primeval-like, ice-age landscape – a landscape that has scarcely changed since. Here you can paddle a canoe for days along the many rivers and lakes. You can take a fishing rod with you to catch the large trout that can be found here, and if you keep your eyes open to the beautiful landscape, you will be rewarded with a memorable trip. The area has been used by the Sámi people for centuries, but retains its unspoilt character. Together with adjoining protected areas in Sweden, the Femundsmarka National Park forms part of one of the largest continuous, undisturbed wilderness areas in southern Scandinavia. It is also an area that provides a habitat for a number of rare and vulnerable animal and bird species. Autumnal colouring in Femundsmarka (BL) 4° 5° Femundsmarka National Park Femundsmarka National Park Canoeing (BL) Evening light on upland downy birches (BL) ENJOY THE SCENERY About half the visitors to the Femundsmarka National Take a trip! Park go canoeing or fishing. The area is also spectacular For hunting, berry-picking, skiing or hiking, Femunds- for hiking - whether you want to go hunting or berry- marka offers plenty of options. During the summer picking, or just enjoy the scenery. -

Nasjonalt Referansesystem for Landskap Beskrivelse Av Norges 45 Landskapsregioner Oskar Puschmann
Nasjonalt referansesystem for landskap Beskrivelse av Norges 45 landskapsregioner Oskar Puschmann NIJOS rapport 10/2005 Nasjonalt referansesystem for landskap - beskrivelse av Norges 45 landskapsregioner av Oskar Puschmann Forsidefoto: Oskar Puschmann Heimlandet på Røst, Røst kommune, Nordland. Landskapsregion 30 Nordlandsverran. NIJOS rapport 10/2005 Tittel: Nasjonalt referansesystem for landskap. NIJOS nummer: Beskrivelse av Norges 45 landskapsregioner 10 /2005 Forfatter(e): ISBN nummer: Oskar Puschmann 82-7464-355-0 Oppdragsgiver: Dato: 13. des. 2005 Landbruks- og matdepartementet Prosjekt/Program: Nasjonalt referansesystem for landskap (RSL) Relatert informasjon/Andre publikasjoner i utvalg fra prosjektet: - Puschmann, O., Reid, S.J., Fjellstad, W., Hofsten, J. & Dramstad, W. 2004. Tilstandsbeskrivelse av norske jordbruksregioner ved bruk av statistikk. NIJOS-rapport 17/04. - Puschmann, O. 1998. Nasjonalt referansesystem for landskap. Bruk av ulike kilder som grunnlag for beskrivelse av underregioner. NIJOS-rapport 12/98. - Kamfjord, G., Lykkja, H. & Puschmann, O. 1997. Landskapet og reiselivsproduktet. NIJOS-rapport 04/97. - Elgersma, A. 1996. ”Norske landskapsregioner. Kart M 1: 2 mill.” Norsk institutt for jord og skogkartlegging.” Utdrag: Siden 1989 har NIJOS arbeidet med utviklingen av et nasjonalt referansesystem for landskap. Dette er et hierarkisk system med inndeling av landet i 45 landskapsregioner og 444 underregioner. Med utgangspunkt i underregionene kan det videre avgrenses i landskapsområder på lokalt nivå. I denne rapporten gis en innføring i metoden som ligger til grunn. Videre inneholder rapporten en beskrivelse, samt visualisering med kart og bilder, av hver av de 45 landskapsregionene. Beskrivelsene skildrer seks ulike landskapskomponenter; landskapets hovedform, landskapets småformer, vann og vassdrag, vegetasjon, jordbruksmark og bebyggelse og tekniske anlegg. I tillegg følger en beskrivelse av regionens samlede landskapskarakter, og her er det også belyst enkelte særegne regionale kvaliteter, problemområder eller utviklingstrender. -

Fiskeguide Roasten 7 12 Stortjønn 5 4
Røros 3 ENGERDAL Røvola Nedre Fiskeguide Roasten 7 12 Stortjønn 5 4 2 Øvre 8 1 Femund Ø, R, H, S, A, G 65 Femundshytta Røvoltjønnan Roasten Storfisk- 2 Røvoltjerna Ø, R, A 6 tjønnan Buhøgda Litle 783 3 Nedre Røa Ø, H, G 4 Vonsjøen 11 Haugen gård 4 Skogtjern Ø, R Litlevonsjø- Det gjøres oppmerksom på at det er en Grøtåa vola Korstjern Ø, R mengde tjern, åer og bekker med fine fiskebe- stander som ikke er omtalt eller kommer fram i 9 5 Roastsjøene Ø, R, H, A, G fiskekartet. Ugland ITs turkartserie i målestokk Grøtådalsætra 10 788 6 Grøtåa Ø 1:50 000, kartblad Femund nord, Femund sør Kuvolsætra Vonsjøen 7 Øvre Røa Ø, R, H, G, A og Engerdal dekker hele kommunen og kan 1416 Grøtådalen 8 Storfisktjern Ø, R, H, G, A anbefales. Flata Stor-Svuku 9 Kratla Ø, R, A Ø = Ørret Svukuris- 10 Store Vonsjøen Ø, R, A, G R = Røye Jonasvollen Rønsjø- Svukuriset Litl-Svuku 11 Lille Vonsjøen R, A H = Harr floan ruten 12 Rogen Ø, R, H, A, G S = Sik Revlinga 13 Rønsjøen Ø, R A = Abbor 13 Revling- 888 Rønsjøen G = Gjedde 14 81 14 Revlingsjøen Ø, R sjøan Røa M = Mort 1 Sandtjønn- 15 Lille Grøvelsjøen Ø 16 Elgåsjøen Ø, R, A vola 17 Volsjøen Ø, R Korneset Øyan Os Sylen 18 Hansbekktjern Ø, R Røsten Elgå Ryvang 82 19 Djupsjøen Ø, R, A Egga Grøvelsjøen Sylens private område Litlå- Kvernvika Elgåhogna 20 Båthussjøen Ø, A 1460 Tolga vollen Solli 21 Storbekktjerna Ø Litlgrøvelsjø- Sals- 22 Flåtesjøen Ø Gjotsætra Johnsgård Sjøli Dalset 15 floan fjellet 23 Skogtjern Ø,R 52 Tjønnodd- Buvika 16 Digerhogna 24 Storsjøen Ø,R,A Langsjøen vola 17 25 Hammersjøen Ø,R,A 18 Volsjøen -

Skadde Fiskebestander I Norske Innsjøer Fra Rundt 1990 Til 2006
169 Endringer i areal med forsurings- skadde fiskebestander i norske innsjøer fra rundt 1990 til 2006 Trygve Hesthagen Gunnel Østborg Rapport nr 123 Naturens Tålegrenser Programmet Naturens Tålegrenser ble satt igang i 1989 i regi av Miljøverndepartementet. Programmet skal blant annet gi innspill til arbeidet med Nordisk Handlingsplan mot Luftforurens- ninger og til aktiviteter under Konvensjonen for Langtransporterte Grensoverskridende Luftforurens- ninger (Genevekonvensjonen). I arbeidet under Genevekonvensjonen er det vedtatt at kritiske belastningsgrenser skal legges til grunn ved utarbeidelse av nye avtaler om utslippsbegrensning av svovel, nitrogen og hydrokarboner. Miljøverndepartementet har det overordnete ansvar for programmet, mens ansvaret for den faglige oppfølgingen er overlatt en arbeidsgruppe bestående av representanter fra Direktoratet for naturforvaltning (DN) og Statens forurensningstilsyn (SFT). Arbeidsgruppen har for tiden følgende sammensetning: Tor Johannessen - SFT Else Løbersli - DN Steinar Sandøy – DN Henvendelse vedrørende programmet kan rettes til: Direktoratet for naturforvaltning 7485 Trondheim Tel: 73 58 05 00 eller Statens forurensningstilsyn Postboks 8100 Dep 0032 Oslo 1 Tel: 22 57 34 00 Norsk institutt for naturforskning Endringer i areal med forsurings- skadde fiskebestander i norske innsjøer fra rundt 1990 til 2006 Trygve Hesthagen Gunnel Østborg NINA Rapport 169 Endringer i areal med forsuringsskadde fiskebestander i norske innsjøer fra rundt 1990 til 2006. - NINA Rapport 169. 114 s. Trondheim, desember 2008 ISSN: 1504-3312 ISBN: 82-426-1724-4 RETTIGHETSHAVER © Norsk institutt for naturforskning Publikasjonen kan siteres fritt med kildeangivelse TILGJENGELIGHET Åpen PUBLISERINGSTYPE Digitalt dokument (pdf) REDAKSJON Odd Terje Sandlund KVALITETSSIKRET AV Odd Terje Sandlund ANSVARLIG SIGNATUR Odd Terje Sandlund (sign.) OPPDRAGSGIVER(E) Direktoratet for naturforvaltning KONTAKTPERSON(ER) HOS OPPDRAGSGIVER Else Løbersli FORSIDEBILDE Videdalsvassdraget i Rogaland. -

Nazewnictwo Geograficzne Ś Wiata
NAZEWNICTWO GEOGRAFICZNE NAZEWNICTWO GEOGRAFICZNE Ś WIATA Zeszyt 12 Ś WIATA 12. Europa – Cz WIATA 12. Europa Europa Część II ęść II IISBNSBN 9978-83-254-0825-178-83-254-0825-1 GŁÓWWNYNY UURZRZĄD GGEODEZJIEODEZJI I KKARTOGRAFIIARTOGRAFII OOkladka.inddkladka.indd 226767 66/10/10/10/10 110:13:170:13:17 PPMM KOMISJA STANDARYZACJI NAZW GEOGRAFICZNYCH POZA GRANICAMI RZECZYPOSPOLITEJ POLSKIEJ przy Głównym Geodecie Kraju NAZEWNICTWO GEOGRAFICZNE ŚWIATA Zeszyt 12 Europa Część II GŁÓWNY URZĄD GEODEZJI I KARTOGRAFII Warszawa 2010 EEuropa2.indburopa2.indb 1 66/10/10/10/10 9:49:219:49:21 PPMM KOMISJA STANDARYZACJI NAZW GEOGRAFICZNYCH POZA GRANICAMI RZECZYPOSPOLITEJ POLSKIEJ przy Głównym Geodecie Kraju Waldemar Rudnicki (przewodniczący), Andrzej Markowski (zastępca przewodniczącego), Maciej Zych (zastępca przewodniczącego), Katarzyna Przyszewska (sekretarz); członkowie: Stanisław Alexandrowicz, Andrzej Czerny, Janusz Danecki, Janusz Gołaski, Romuald Huszcza, Sabina Kacieszczenko, Dariusz Kalisiewicz, Artur Karp, Zbigniew Obidowski , Jerzy Ostrowski, Jarosław Pietrow, Jerzy Pietruszka, Andrzej Pisowicz, Bogumiła Więcław, Ewa Wolnicz-Pawłowska, Bogusław R. Zagórski Opracowanie Elżbieta Chudorlińska (Belgia, Holandia, Irlandia, Szwajcaria, Szwecja, Wielka Brytania), Andrzej Czerny (Austria, Niemcy), Jarosław Pietrow (Dania, Finlandia, Islandia, Norwegia, Szwecja), Agnieszka Pietrzak (Hiszpania, Malta, Portugalia, Włochy), Maksymilian Skotnicki (Francja), Maciej Zych (Andora, Francja, Gibraltar, Guernsey, Holandia, Jan Mayen, Jersey, Liechtenstein, Luksemburg, -
Aurebestanden I Roastensjøene
Campus Evenstad Skog og utmarksfag Marius Hassve Aurebestanden i Roastensjøene - Villfisk og settefisk Bacheloroppgave i Utmarksforvaltning 2010 Utlånsklausul: Nei Ja. Antall år Høgskolen i Hedmark Høgskoleadministrasjonen Postadresse: 2418 Elverum Besøksadresse: Lærerskolealléen 1 Telefon +47 62 43 00 00 Telefaks +47 62 43 00 01 E-post [email protected] Org. nr 974 251 760 www.hihm.no Sammendrag Hassve, M. (2010) Aurebestanden i Roastensjøene – Villfisk og settefisk. 44 sider inkludert vedlegg. Målsetningen med denne oppgaven har vært å få en oversikt over aurebestanden i Nedre og Øvre Roasten i Femundsmarka Nasjonalpark, med hovedvekt på forholdet mellom villfisk og settefisk, og derav metodikken vedrørende det å skille mellom den naturlige og utsatte auren. Ved hjelp av prøvefiske med garn har aurebestanden i Nedre og Øvre Roasten blitt klassifisert som tynn med fisk av middels størrelse. Auren er av god kvalitet, og vekstmønsteret samt den gjennomsnittlige lengden for gytemodne hunner, kan tyde på at auren ikke er næringsbegrenset. Ut i fra undersøkelsene ser det ut til å være gode gytemuligheter, og selv om det med elektrisk fiskeapparat og lystringsfiske ikke ble fanget rekrutter yngre enn 1 år, synes det, med bakgrunn i aldersfordelingen, å være god naturlig rekruttering. Villfisk og settefisk er skilt ved å se på første års vekst ved hjelp av strukturene otolitter og skjell. Aure, der skjell har stor sklerittavstand og lang første vekstsesong, har blitt klassifisert som settefisk, mens aure med tette skleritter, og kort første vekstsesong har blitt klassifisert som villfisk. Resultatene viser at villfisk har en signifikant dårligere vekst første vekstsesong enn settefisk. Settefisk fra Snerta settefiskanlegg har blitt benyttet som sammenligningsgrunnlag.