Cerebellar Development and Neurogenesis in Zebrafish
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ultrastructural Study of the Granule Cell Domain of the Cochlear Nucleus in Rats: Mossy Fiber Endings and Their Targets
THE JOURNAL OF COMPARATIVE NEUROLOGY 369~345-360 ( 1996) Ultrastructural Study of the Granule Cell Domain of the Cochlear Nucleus in Rats: Mossy Fiber Endings and Their Targets DIANA L. WEEDMAN, TAN PONGSTAPORN, AND DAVID K. RYUGO Center for Hearing Sciences, Departments of Otolaryngoloby-Head and Neck Surgery and Neuroscience, Johns Hopkins University School of Medicine, Baltimore, Maryland 2 1205 ABSTRACT The principal projection neurons of the cochlear nucleus receive the bulk of their input from the auditory nerve. These projection neurons reside in the core of the nucleus and are surrounded by an external shell, which is called the granule cell domain. Interneurons of the cochlear granule cell domain are the target for nonprimary auditory inputs, including projections from the superior olivary complex, inferior colliculus, and auditory cortex. The granule cell domain also receives projections from the cuneate and trigeminal nuclei, which are first-order nuclei of the somatosensory system. The cellular targets of the nonprimary projections are mostly unknown due to a lack of information regarding postsynaptic profiles in the granule cell areas. In the present paper, we examined the synaptic relationships between a heterogeneous class of large synaptic terminals called mossy fibers and their targets within subdivisions of the granule cell domain known as the lamina and superficial layer. By using light and electron microscopic methods in these subdivisions, we provide evidence for three different neuron classes that receive input from the mossy fibers: granule cells, unipolar brush cells, and a previously undescribed class called chestnut cells. The distinct synaptic relations between mossy fibers and members of each neuron class further imply fundamentally separate roles for processing acoustic signals. -

Consensus Paper: Cerebellar Development
Cerebellum DOI 10.1007/s12311-015-0724-2 CONSENSUS PAPER Consensus Paper: Cerebellar Development Ketty Leto1,2 & Marife Arancillo3 & Esther B. E. Becker4 & Annalisa Buffo1,2 & Chin Chiang5 & Baojin Ding6 & William B. Dobyns 7,8 & Isabelle Dusart9,10 & Parthiv Haldipur7 & Mary E. Hatten11 & Mikio Hoshino12 & Alexandra L. Joyner13 & Masanobu Kano14 & Daniel L. Kilpatrick6 & Noriyuki Koibuchi15 & Silvia Marino16 & Salvador Martinez17 & Kathleen J. Millen7 & Thomas O. Millner16 & Takaki Miyata18 & Elena Parmigiani1,2 & Karl Schilling19 & Gabriella Sekerková20 & Roy V. Sillitoe3 & Constantino Sotelo21 & Naofumi Uesaka14 & Annika Wefers 22 & Richard J. T. Wingate23 & Richard Hawkes24 # The Author(s) 2015. This article is published with open access at Springerlink.com Abstract The development of the mammalian cerebellum is processes of cerebellar ontogenesis, highlighting the neuro- orchestrated by both cell-autonomous programs and inductive genic strategies used by developing progenitors, the genetic environmental influences. Here, we describe the main programs involved in cell fate specification, the progressive * Ketty Leto 10 Centre National de la Recherche Scientifique, CNRS, UMR8246, [email protected] INSERM U1130, Neuroscience Paris Seine, France, 75005 Paris, France 11 Laboratory of Developmental Neurobiology, The Rockefeller 1 Department of Neuroscience Rita Levi Montalcini, University of University, New York, NY 10065, USA Turin, via Cherasco 15, 10026 Turin, Italy 12 Department of Biochemistry and Cellular Biology, National Institute -

Purkinje Cell Migration Disorder By
CEREBELLAR CORTICOGENESIS IN THE LYSOSOMAL ACID PHOSPHATASE (ACP2) MUTANT MICE: PURKINJE CELL MIGRATION DISORDER BY NILOUFAR ASHTARI A Thesis Submitted to the Faculty of Graduate Studies of The University of Manitoba in Partial Fulfilment of the Requirements for the Degree of MASTER OF SCIENCE Department of Human Anatomy and Cell Science University of Manitoba Winnipeg, Manitoba Copyright © 2017 by Niloufar Ashtari 1 Abstract In a mutant mouse called nax as the result of mutation in Lysosomal Acid phosphatase (Acp2), layers of the cerebellar cortex are impaired and monolayer Purkinje cells (Pcs) turn to multi-layered Pcs that ectopically invade the molecular layer. We investigated reelin-Dab1 signaling as an important pathway for Pcs migration and monolayer formation in cerebellum. ERK1/2 is a member of mitogen activated kinases family and suggested to be a downstream of reelin signaling. We hypothesize that the establishment of mono-layered Pcs rely on reelin through ERK1/2 pathway. Acp2 mutant mice were used for this study and molecular expression and distribution were assessed by immunohistochemistry, RT-PCR, western blotting, and cell culture. Results suggest that reelin may modulate the ERK1/2 expression, thus lower expression of reelin and higher phosphorylation of Dab1 leads to over expression of the ERK1/2 that causes the Pcs to over migrate and form multilayer in nax cerebellar cortex. i TABLE OF CONTENTS LISTOFABBREVIATIONS………………………………………………..…... IV LIST OF TABLES……………………………………...…………………...….. Vii LIST OF FIGURES…………………………………………………….………. Viii CHAPTER 1: INTRODUCTION…………………………………….………… 1 1.1 Cerebellum ……………………………………………………........………. 1 1.2 Development of Central Nervous System………………………………….. 2 1.3 Development of the cerebellum………………………………….................. 3 1.4 Specification of cerebellar germinal zones…………………………………. -

The Cerebellum in Sagittal Plane-Anatomic-MR Correlation: 2
667 The Cerebellum in Sagittal Plane-Anatomic-MR Correlation: 2. The Cerebellar Hemispheres Gary A. Press 1 Thin (5-mm) sagittal high-field (1 .5-T) MR images of the cerebellar hemispheres James Murakami2 display (1) the superior, middle, and inferior cerebellar peduncles; (2) the primary white Eric Courchesne2 matter branches to the hemispheric lobules including the central, anterior, and posterior Dean P. Berthoty1 quadrangular, superior and inferior semilunar, gracile, biventer, tonsil, and flocculus; Marjorie Grafe3 and (3) several finer secondary white-matter branches to individual folia within the lobules. Surface features of the hemispheres including the deeper fissures (e.g., hori Clayton A. Wiley3 1 zontal, posterolateral, inferior posterior, and inferior anterior) and shallower sulci are John R. Hesselink best delineated on T1-weighted (short TRfshort TE) and T2-weighted (long TR/Iong TE) sequences, which provide greatest contrast between CSF and parenchyma. Correlation of MR studies of three brain specimens and 11 normal volunteers with microtome sections of the anatomic specimens provides criteria for identifying confidently these structures on routine clinical MR. MR should be useful in identifying, localizing, and quantifying cerebellar disease in patients with clinical deficits. The major anatomic structures of the cerebellar vermis are described in a companion article [1). This communication discusses the topographic relationships of the cerebellar hemispheres as seen in the sagittal plane and correlates microtome sections with MR images. Materials, Subjects, and Methods The preparation of the anatomic specimens, MR equipment, specimen and normal volunteer scanning protocols, methods of identifying specific anatomic structures, and system of This article appears in the JulyI August 1989 issue of AJNR and the October 1989 issue of anatomic nomenclature are described in our companion article [1]. -

Staining of Cerebellar Cortex Granular Layer Interneurons with Natural Dye of Madder Anneh Mohammad Gharravi
Gharravi Cerebellum & Ataxias (2016) 3:12 DOI 10.1186/s40673-016-0050-6 RESEARCH Open Access Staining of cerebellar cortex granular layer interneurons with natural dye of Madder Anneh Mohammad Gharravi Abstract Background: The objective of the present study was an investigation of root Rubia Tinctorum (Madder) as a natural dye to identification of granular layer interneurons of the rat cerebellum. Methods: Seven to ten micrometre sections were collected from the cerebellum and stained only with Madder for 2, 24 and 48 h. Other sections were stained with Madder then with hematoxyllin, cresyl violet, eosin, light green. Microscopic identification of cells was performed based on cell morphology, reaction and binding of with the dye. All data were expressed as mean ± SD in and significance was set at p ≤0.05. Results: Madder with alum as mordant resulted a deep red staining of interneurons. Unipolar brush cells (UBCs) were observed with a cell body diameter intermediate between granule and Golgi cells in the superficial layer of the granular layer. Golgi cells were identified almost as large as Purkinje cells with irregular rounded or polygonal morphology. Lugaro cells were observed as spindle-shaped cells adjacent to Purkinje layer. Conclusion: Results of the present study showed that mader could stain granular layer interneurons in cerebellum cortex of rat. Keywords: Madder, Cerebellum, Unipolar brush cells, Lugaro cell, Golgi neurons Background with all cerebellar cortical neurons and fibers and they Histologically, the cerebellar cortex is divided into three function as inhibitory interneurons. These spindle-shaped layers: the molecular, the Purkinje and the granular cells locate just underneath the Purkinje cell layer are layers. -

Occurrence of Long-Term Depression in the Cerebellar Flocculus During Adaptation of Optokinetic Response Takuma Inoshita, Tomoo Hirano*
SHORT REPORT Occurrence of long-term depression in the cerebellar flocculus during adaptation of optokinetic response Takuma Inoshita, Tomoo Hirano* Department of Biophysics, Graduate School of Science, Kyoto University, Sakyo-ku, Japan Abstract Long-term depression (LTD) at parallel fiber (PF) to Purkinje cell (PC) synapses has been considered as a main cellular mechanism for motor learning. However, the necessity of LTD for motor learning was challenged by demonstration of normal motor learning in the LTD-defective animals. Here, we addressed possible involvement of LTD in motor learning by examining whether LTD occurs during motor learning in the wild-type mice. As a model of motor learning, adaptation of optokinetic response (OKR) was used. OKR is a type of reflex eye movement to suppress blur of visual image during animal motion. OKR shows adaptive change during continuous optokinetic stimulation, which is regulated by the cerebellar flocculus. After OKR adaptation, amplitudes of quantal excitatory postsynaptic currents at PF-PC synapses were decreased, and induction of LTD was suppressed in the flocculus. These results suggest that LTD occurs at PF-PC synapses during OKR adaptation. DOI: https://doi.org/10.7554/eLife.36209.001 Introduction The cerebellum plays a critical role in motor learning, and a type of synaptic plasticity long-term *For correspondence: depression (LTD) at parallel fiber (PF) to Purkinje cell (PC) synapses has been considered as a primary [email protected]. cellular mechanism for motor learning (Ito, 1989; Hirano, 2013). However, the hypothesis that LTD ac.jp is indispensable for motor learning was challenged by demonstration of normal motor learning in Competing interests: The rats in which LTD was suppressed pharmacologically or in the LTD-deficient transgenic mice authors declare that no (Welsh et al., 2005; Schonewille et al., 2011). -

FIRST PROOF Cerebellum
Article Number : EONS : 0736 GROSS ANATOMY Cerebellum Cortex The cerebellar cortex is an extensive three-layered sheet with a surface approximately 15 cm laterally THE HUMAN CEREBELLUM (‘‘little brain’’) is a and 180 cm rostrocaudally but densely folded around significant part of the central nervous system both three pairs of nuclei. The cortex is divided into three in size and in neural structure. It occupies approxi- transverse lobes: Anterior and posterior lobes are mately one-tenth of the cranial cavity, sitting astride separated by the primary fissure, and the smaller the brainstem, beneath the occipital cortex, and flocculonodular lobe is separated by the poster- contains more neurons than the whole of the cerebral olateral fissure (Fig. 1). The anterior and posterior cortex. It consists of an extensive cortical sheet, lobes are folded into a number of lobules and further densely folded around three pairs of nuclei. The folded into a series of folia. This transverse organiza- cortex contains only five main neural cell types and is tion is then divided at right angles into broad one of the most regular and uniform structures in the longitudinal regions. The central vermis, named for central nervous system (CNS), with an orthogonal its worm-like appearance, is most obvious in the ‘‘crystalline’’ organization. Major connections are posterior lobe. On either side is the paravermal or made to and from the spinal cord, brainstem, and intermediate cortex, which merges into the lateral sensorimotor areas of the cerebral cortex. hemispheres. The most common causes of damage to the cerebellum are stroke, tumors, or multiple sclerosis. -

Cerebellar Cortical Neuron Responses Evoked from the Spinal Border Cell Tract
Cerebellar cortical neuron responses evoked from the spinal border cell tract. Geborek, Pontus; Spanne, Anton; Bengtsson, Fredrik; Jörntell, Henrik Published in: Frontiers in Neural Circuits DOI: 10.3389/fncir.2013.00157 2013 Link to publication Citation for published version (APA): Geborek, P., Spanne, A., Bengtsson, F., & Jörntell, H. (2013). Cerebellar cortical neuron responses evoked from the spinal border cell tract. Frontiers in Neural Circuits, 7, [157]. https://doi.org/10.3389/fncir.2013.00157 Total number of authors: 4 General rights Unless other specific re-use rights are stated the following general rights apply: Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal Read more about Creative commons licenses: https://creativecommons.org/licenses/ Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. LUND UNIVERSITY PO Box 117 221 00 Lund +46 46-222 00 00 Download date: 04. Oct. 2021 ORIGINAL RESEARCH ARTICLE published: 08 October 2013 NEURAL CIRCUITS doi: 10.3389/fncir.2013.00157 Cerebellar cortical neuron responses evoked from the spinal border cell tract Pontus Geborek, Anton Spanne, Fredrik Bengtsson and Henrik Jörntell* Neural Basis of Sensorimotor Control, Department of Experimental Medical Science, Lund University, Lund, Sweden Edited by: Spinocerebellar systems are likely to be crucial for cerebellar hallmark functions such Egidio D’Angelo, University of Pavia, as coordination. -

Forward Signaling by Unipolar Brush Cells in the Mouse Cerebellum
Cerebellum DOI 10.1007/s12311-015-0693-5 ORIGINAL PAPER Forward Signaling by Unipolar Brush Cells in the Mouse Cerebellum Stijn van Dorp1 & Chris I. De Zeeuw 1,2 # The Author(s) 2015. This article is published with open access at Springerlink.com Abstract Unipolar brush cells (UBCs) are glutamatergic in- Introduction terneurons prominently present in the granular layer of the vestibulocerebellum. UBCs engage in extensive synaptic con- Forward processing, in particular the absence of recurrent tact with a single presynaptic mossy fiber and signal to down- excitation, is a defining feature of cerebellar architecture stream granule cells through an elaborate network of mossy and computation. In the granular layer of the fiber-like axons. Ultrastructural examinations and electro- vestibulocerebellum, unipolar brush cells (UBCs) provide physiological recordings in organotypic slice cultures have a powerful forward excitatory action onto granule cells indicated that UBCs target not only granule cells but also other through a cortex-intrinsic network of mossy fiber-like UBCs, thus forming chains of two or perhaps more intercon- axons [1, 2]. UBCs are characterized by an elaborate nected UBCs. In this report, we show recordings of spontane- brush-like dendrite (Fig. 1a) that forms an unusually exten- ous and evoked (di)synaptic events in granule cells and UBCs sive synaptic contact with a single presynaptic mossy fiber in fresh cerebellar slices from juvenile mice (5–7 weeks). The rosette [3]. This highly specialized configuration has been patterns of arrival of synaptic events were consistent with the proposed to facilitate prolonged entrapment of glutamate presence of a presynaptic UBC, and recordings from UBCs in the synaptic cleft, underlying complex temporal trans- displayed spontaneous protracted synaptic events characteris- formations of incoming mossy fiber signals [4, 5]. -

…Going One Step Further
…going one step further C20 (1017868) 2 Latin A Encephalon Mesencephalon B Telencephalon 31 Lamina tecti B1 Lobus frontalis 32 Tegmentum mesencephali B2 Lobus temporalis 33 Crus cerebri C Diencephalon 34 Aqueductus mesencephali D Mesencephalon E Metencephalon Metencephalon E1 Cerebellum 35 Cerebellum F Myelencephalon a Vermis G Circulus arteriosus cerebri (Willisii) b Tonsilla c Flocculus Telencephalon d Arbor vitae 1 Lobus frontalis e Ventriculus quartus 2 Lobus parietalis 36 Pons 3 Lobus occipitalis f Pedunculus cerebellaris superior 4 Lobus temporalis g Pedunculus cerebellaris medius 5 Sulcus centralis h Pedunculus cerebellaris inferior 6 Gyrus precentralis 7 Gyrus postcentralis Myelencephalon 8 Bulbus olfactorius 37 Medulla oblongata 9 Commissura anterior 38 Oliva 10 Corpus callosum 39 Pyramis a Genu 40 N. cervicalis I. (C1) b Truncus ® c Splenium Nervi craniales d Rostrum I N. olfactorius 11 Septum pellucidum II N. opticus 12 Fornix III N. oculomotorius 13 Commissura posterior IV N. trochlearis 14 Insula V N. trigeminus 15 Capsula interna VI N. abducens 16 Ventriculus lateralis VII N. facialis e Cornu frontale VIII N. vestibulocochlearis f Pars centralis IX N. glossopharyngeus g Cornu occipitale X N. vagus h Cornu temporale XI N. accessorius 17 V. thalamostriata XII N. hypoglossus 18 Hippocampus Circulus arteriosus cerebri (Willisii) Diencephalon 1 A. cerebri anterior 19 Thalamus 2 A. communicans anterior 20 Sulcus hypothalamicus 3 A. carotis interna 21 Hypothalamus 4 A. cerebri media 22 Adhesio interthalamica 5 A. communicans posterior 23 Glandula pinealis 6 A. cerebri posterior 24 Corpus mammillare sinistrum 7 A. superior cerebelli 25 Hypophysis 8 A. basilaris 26 Ventriculus tertius 9 Aa. pontis 10 A. -

The Developmental Genetics of the Cerebellum and the Genetic Bases of Known Cerebellar Disorders
THE DEVELOPMENTAL GENETICS OF THE CEREBELLUM AND THE GENETIC BASES OF KNOWN CEREBELLAR DISORDERS A Review of Developmental Genetics of the Cerebellum and the Genetic Bases of known Cerebellar Disorders Olakada Favour Adamba 17/MHS01/254 Department of Medicine and Health Sciences College of Medicine and Health Sciences Afe Babalola University ANA 303: Neuroanatomy July, 2020 The Developmental Genetics of Cerebellum and the Genetic Bases of known Cerebellar Disorders: A Literature Review Olakada Favour Adamba1 Abstract The internal structure of the cerebellum is an intriguing paradox; its cytoarchitecture is relatively simple compared to the connections between its neurons, which are wired into a complex array of gene expression domains and functional circuits. The genetic research of cerebellar development has provided a great deal of information about the molecular events directing the formation of the cerebellum. The developmental mechanisms that coordinate the establishment of cerebellar structure and circuitry provide a powerful model for understanding how a functional brain develops and its significance in cerebellar disorders and diseases. The cellular makeup of the cerebellum is derived from two primary germinal matrices (the ventricular zone and a specialized germinal matrix called the rhombic lip). Each matrix/zone expresses a specific set of genes that establish the cell lineages within the cerebellar anlage. Then, cohorts of differentiated projection neurons and interneuron progenitors migrate into the developing cerebellum. thereafter, a number of remarkable patterning events occur. Altogether, structural and molecular organisations are thought to support the proper connectivity between incoming afferent projections and their target cells. Key words: Cerebellum, circuitry, genetic, development, disorders. I. Introduction The cerebellum (‘little brain’) resides at the anterior end of the hindbrain and is classically defined by its role in sensory-motor processing (Buckner, 2013). -
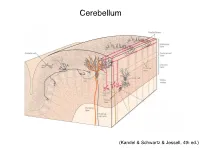
Cerebellum and Activation of the Cerebellum IO ST During Nonmotor Tasks
Cerebellum (Kandel & Schwartz & Jessell, 4th ed.) Granule186 cells are irreducibly smallChapter (6-8 7 μm) stellate cell basket cell outer synaptic layer PC rs cap PC layer grc Go inner 6 μm synaptic layer Figure 7.15 Largest cerebellar neuron occupies more than a 1,000-fold greater volume than smallest neuron. Thin section (~1 μ m) through monkey cerebellar cortex. Purkinje cell body (PC) and nucleus are far larger than those of granule cell (grc). The latter cluster to leave space for mossy fiber terminals to form glomeruli with grc dendritic claws and space for Golgi cells (Go). Note rich network of capillaries (cap). Fine, scat- tered dots are mitochondria. Courtesy of E. Mugnaini. brain ’ s most numerous neuron, this small cost grows large (see also chapter 13). Much of the inner synaptic layer is occupied by the large axon terminals of mossy fibers (figures 7.1 and 7.16). A terminal interlaces with multiple (~15) dendritic claws, each from a different but neighboring granule cell, and forms a complex knot (glomerulus ), nearly as large as a granule cell body (figures 7.1 and 7.16). The mossy fiber axon fires at an unusually high mean rate (up to 200 Hz) and is therefore among the brain’ s thickest (figure 4.6). To match the axon’ s high rate, a terminal expresses 150 active zones, 10 per postsynaptic granule cell (figure 7.16). These sites are capable of driving … and most expensive. 190 Chapter 7 10 4 8 3 9 19 10 10 × 6 × 2 2 4 ATP/s/cell ATP/s/cell ATP/s/m 1 2 0 0 astrocyte astrocyte Golgi cell Golgi cell basket cell basket cell stellate cell stellate cell granule cell granule cell mossy fiber mossy fiber Purkinje cell Purkinje Purkinje cell Purkinje Bergman glia Bergman glia climbing fiber climbing fiber Figure 7.18 Energy costs by cell type.