On the Function of the Floccular Complex of the Vertebrate Cerebellum: Implications in Paleoneuroanatomy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Cerebellum in Sagittal Plane-Anatomic-MR Correlation: 2
667 The Cerebellum in Sagittal Plane-Anatomic-MR Correlation: 2. The Cerebellar Hemispheres Gary A. Press 1 Thin (5-mm) sagittal high-field (1 .5-T) MR images of the cerebellar hemispheres James Murakami2 display (1) the superior, middle, and inferior cerebellar peduncles; (2) the primary white Eric Courchesne2 matter branches to the hemispheric lobules including the central, anterior, and posterior Dean P. Berthoty1 quadrangular, superior and inferior semilunar, gracile, biventer, tonsil, and flocculus; Marjorie Grafe3 and (3) several finer secondary white-matter branches to individual folia within the lobules. Surface features of the hemispheres including the deeper fissures (e.g., hori Clayton A. Wiley3 1 zontal, posterolateral, inferior posterior, and inferior anterior) and shallower sulci are John R. Hesselink best delineated on T1-weighted (short TRfshort TE) and T2-weighted (long TR/Iong TE) sequences, which provide greatest contrast between CSF and parenchyma. Correlation of MR studies of three brain specimens and 11 normal volunteers with microtome sections of the anatomic specimens provides criteria for identifying confidently these structures on routine clinical MR. MR should be useful in identifying, localizing, and quantifying cerebellar disease in patients with clinical deficits. The major anatomic structures of the cerebellar vermis are described in a companion article [1). This communication discusses the topographic relationships of the cerebellar hemispheres as seen in the sagittal plane and correlates microtome sections with MR images. Materials, Subjects, and Methods The preparation of the anatomic specimens, MR equipment, specimen and normal volunteer scanning protocols, methods of identifying specific anatomic structures, and system of This article appears in the JulyI August 1989 issue of AJNR and the October 1989 issue of anatomic nomenclature are described in our companion article [1]. -

Occurrence of Long-Term Depression in the Cerebellar Flocculus During Adaptation of Optokinetic Response Takuma Inoshita, Tomoo Hirano*
SHORT REPORT Occurrence of long-term depression in the cerebellar flocculus during adaptation of optokinetic response Takuma Inoshita, Tomoo Hirano* Department of Biophysics, Graduate School of Science, Kyoto University, Sakyo-ku, Japan Abstract Long-term depression (LTD) at parallel fiber (PF) to Purkinje cell (PC) synapses has been considered as a main cellular mechanism for motor learning. However, the necessity of LTD for motor learning was challenged by demonstration of normal motor learning in the LTD-defective animals. Here, we addressed possible involvement of LTD in motor learning by examining whether LTD occurs during motor learning in the wild-type mice. As a model of motor learning, adaptation of optokinetic response (OKR) was used. OKR is a type of reflex eye movement to suppress blur of visual image during animal motion. OKR shows adaptive change during continuous optokinetic stimulation, which is regulated by the cerebellar flocculus. After OKR adaptation, amplitudes of quantal excitatory postsynaptic currents at PF-PC synapses were decreased, and induction of LTD was suppressed in the flocculus. These results suggest that LTD occurs at PF-PC synapses during OKR adaptation. DOI: https://doi.org/10.7554/eLife.36209.001 Introduction The cerebellum plays a critical role in motor learning, and a type of synaptic plasticity long-term *For correspondence: depression (LTD) at parallel fiber (PF) to Purkinje cell (PC) synapses has been considered as a primary [email protected]. cellular mechanism for motor learning (Ito, 1989; Hirano, 2013). However, the hypothesis that LTD ac.jp is indispensable for motor learning was challenged by demonstration of normal motor learning in Competing interests: The rats in which LTD was suppressed pharmacologically or in the LTD-deficient transgenic mice authors declare that no (Welsh et al., 2005; Schonewille et al., 2011). -

Neuroimaging Evidence Implicating Cerebellum in the Experience of Hypercapnia and Hunger for Air
Neuroimaging evidence implicating cerebellum in the experience of hypercapnia and hunger for air Lawrence M. Parsons*†, Gary Egan‡, Mario Liotti*, Stephen Brannan*, Derek Denton‡, Robert Shade§, Rachael Robillard¶, Lisa Madden§, Bart Abplanalp¶, and Peter T. Fox* *Research Imaging Center, University of Texas Health Science Center, San Antonio, TX 78284; ‡Howard Florey Institute of Experimental Physiology and Medicine, University of Melbourne, Parkville, Victoria 3052, Australia; §Southwest Foundation for Biomedical Research, P. O. Box 760549, San Antonio, TX 78245-0549; and ¶Departments of Psychology and Educational Psychology, University of Texas, Austin, TX 78712 Contributed by Derek Denton, December 20, 2000 Recent neuroimaging and neurological data implicate cerebellum areas (20–23). Anatomical tracer labeling data in rat (24) in nonmotor sensory, cognitive, vegetative, and affective func- indicate that a number of cerebellar areas send and͞or receive tions. The present study assessed cerebellar responses when the projections from the VRG, which contains the structures nec- urge to breathe is stimulated by inhaled CO2. Ventilation changes essary for respiratory rhythm generation (25–30). The cerebellar follow arterial blood partial pressure CO2 changes sensed by the areas connected to the VRG are quadrangular (VI), central medullary ventral respiratory group (VRG) and hypothalamus, (III), lingula (I, II), and inferior semilunar (Crus II) lobules, as entraining changes in midbrain, pons, thalamus, limbic, paralimbic, well as fastigial nucleus, interposed nucleus, and dentate nuclei, and insular regions. Nearly all these areas are known to connect the output nuclei for the vermal, intermediate, and lateral anatomically with the cerebellum. Using positron emission tomog- regions of cerebellum. Moreover, cerebellum has known con- raphy, we measured regional brain blood flow during acute CO2- nectivity to the hypothalamus, thalamus, pons, and midbrain induced breathlessness in humans. -

…Going One Step Further
…going one step further C20 (1017868) 2 Latin A Encephalon Mesencephalon B Telencephalon 31 Lamina tecti B1 Lobus frontalis 32 Tegmentum mesencephali B2 Lobus temporalis 33 Crus cerebri C Diencephalon 34 Aqueductus mesencephali D Mesencephalon E Metencephalon Metencephalon E1 Cerebellum 35 Cerebellum F Myelencephalon a Vermis G Circulus arteriosus cerebri (Willisii) b Tonsilla c Flocculus Telencephalon d Arbor vitae 1 Lobus frontalis e Ventriculus quartus 2 Lobus parietalis 36 Pons 3 Lobus occipitalis f Pedunculus cerebellaris superior 4 Lobus temporalis g Pedunculus cerebellaris medius 5 Sulcus centralis h Pedunculus cerebellaris inferior 6 Gyrus precentralis 7 Gyrus postcentralis Myelencephalon 8 Bulbus olfactorius 37 Medulla oblongata 9 Commissura anterior 38 Oliva 10 Corpus callosum 39 Pyramis a Genu 40 N. cervicalis I. (C1) b Truncus ® c Splenium Nervi craniales d Rostrum I N. olfactorius 11 Septum pellucidum II N. opticus 12 Fornix III N. oculomotorius 13 Commissura posterior IV N. trochlearis 14 Insula V N. trigeminus 15 Capsula interna VI N. abducens 16 Ventriculus lateralis VII N. facialis e Cornu frontale VIII N. vestibulocochlearis f Pars centralis IX N. glossopharyngeus g Cornu occipitale X N. vagus h Cornu temporale XI N. accessorius 17 V. thalamostriata XII N. hypoglossus 18 Hippocampus Circulus arteriosus cerebri (Willisii) Diencephalon 1 A. cerebri anterior 19 Thalamus 2 A. communicans anterior 20 Sulcus hypothalamicus 3 A. carotis interna 21 Hypothalamus 4 A. cerebri media 22 Adhesio interthalamica 5 A. communicans posterior 23 Glandula pinealis 6 A. cerebri posterior 24 Corpus mammillare sinistrum 7 A. superior cerebelli 25 Hypophysis 8 A. basilaris 26 Ventriculus tertius 9 Aa. pontis 10 A. -

Anatomy of the Cerebellum Computational Models of Neural
Anatomy of the Cerebellum Computational Models of neural Systems Lecture 2.1 David S. Touretzky September, 2015 First Look cerebellum 09/09/15 Computational Models of Neural Systems 2 Lateral View 09/09/15 Computational Models of Neural Systems 3 Ventral View 09/09/15 Computational Models of Neural Systems 4 Basic Facts About the Cerebellum ● Latin for ªlittle brainº. ● An older brain area, with a simple, regular architecture. ● Makes up 10% of brain volume, but contains over 50% of the brain©s neurons and 4X the neurons of the cerebral cortex. ● Huge fan-in: 40X as many axons enter the cerebellum as exit from it. ● Necessary for smooth, accurate performance of motor actions. ● Example: moving your arm rapidly in a circle. – Involves many muscles in the arm, trunk, and legs. ● People can still move without a cerebellum, but their actions will not be coordinated. There can be overshoots and oscillations. 09/09/15 Computational Models of Neural Systems 5 Cortical Projections to Cerebellum From Strick et al., Annual review of Neuroscience (2009), adapted from Glickstein et al. (1985) J. Comparative Neurology 09/09/15 Computational Models of Neural Systems 6 Three Cerebellar Lobes ● Anterior (divided into 3 lobules) ● Posterior (divided into 6 lobules) ● Flocculonodular 09/09/15 Computational Models of Neural Systems 7 10 Lobules Lingula, Central, Culmen, Declive, Folium, Tuber, Pyramis, Uvula, Tonsil, Flocculonodular 09/09/15 Computational Models of Neural Systems 8 8 of the 10 Lobules 1. Lingula 2. Central Lobule 3. Culmen 4. Declive 5. -
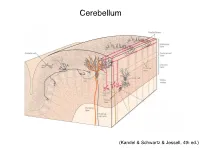
Cerebellum and Activation of the Cerebellum IO ST During Nonmotor Tasks
Cerebellum (Kandel & Schwartz & Jessell, 4th ed.) Granule186 cells are irreducibly smallChapter (6-8 7 μm) stellate cell basket cell outer synaptic layer PC rs cap PC layer grc Go inner 6 μm synaptic layer Figure 7.15 Largest cerebellar neuron occupies more than a 1,000-fold greater volume than smallest neuron. Thin section (~1 μ m) through monkey cerebellar cortex. Purkinje cell body (PC) and nucleus are far larger than those of granule cell (grc). The latter cluster to leave space for mossy fiber terminals to form glomeruli with grc dendritic claws and space for Golgi cells (Go). Note rich network of capillaries (cap). Fine, scat- tered dots are mitochondria. Courtesy of E. Mugnaini. brain ’ s most numerous neuron, this small cost grows large (see also chapter 13). Much of the inner synaptic layer is occupied by the large axon terminals of mossy fibers (figures 7.1 and 7.16). A terminal interlaces with multiple (~15) dendritic claws, each from a different but neighboring granule cell, and forms a complex knot (glomerulus ), nearly as large as a granule cell body (figures 7.1 and 7.16). The mossy fiber axon fires at an unusually high mean rate (up to 200 Hz) and is therefore among the brain’ s thickest (figure 4.6). To match the axon’ s high rate, a terminal expresses 150 active zones, 10 per postsynaptic granule cell (figure 7.16). These sites are capable of driving … and most expensive. 190 Chapter 7 10 4 8 3 9 19 10 10 × 6 × 2 2 4 ATP/s/cell ATP/s/cell ATP/s/m 1 2 0 0 astrocyte astrocyte Golgi cell Golgi cell basket cell basket cell stellate cell stellate cell granule cell granule cell mossy fiber mossy fiber Purkinje cell Purkinje Purkinje cell Purkinje Bergman glia Bergman glia climbing fiber climbing fiber Figure 7.18 Energy costs by cell type. -

Neuroimaging Evidence Implicating Cerebellum in Support of Sensory Cognitive Processes Associated with Thirst
Neuroimaging evidence implicating cerebellum in support of sensory͞cognitive processes associated with thirst Lawrence M. Parsons*†, Derek Denton‡, Gary Egan‡, Michael McKinley‡, Robert Shade‡, Jack Lancaster*, and Peter T. Fox* *Research Imaging Center, Medical School, University of Texas Health Science Center at San Antonio, Floyd Curl Drive, San Antonio, TX 78284; ‡Howard Florey Institute of Experimental Physiology and Medicine, University of Melbourne, Parkville, Victoria 3052, Australia; and §Southwest Foundation for Biomedical Research, P.O. Box 760549, San Antonio, TX 78245-0549 Contributed by Derek A. Denton, December 13, 1999 Recent studies implicate the cerebellum, long considered strictly a and tactile discrimination, kinesthetic sensation, and working motor control structure, in cognitive, sensory, and affective phe- memory, among other processes (6–12). It has been proposed nomenon. The cerebellum, a phylogenetically ancient structure, that the lateral cerebellum may be activated during several has reciprocal ancient connections to the hypothalamus, a struc- motor, perceptual, and cognitive processes specifically because ture important in vegetative functions. The present study investi- of the requirement to monitor and adjust the acquisition of gated whether the cerebellum was involved in vegetative func- sensory data (2, 13). Furthermore, there are reports suggesting tions and the primal emotions engendered by them. Using positron the involvement of posterior vermal cerebellum in affect (14, 15). emission tomography, we examined the effects on the cerebellum The findings implicating the cerebellum in sensory processing of the rise of plasma sodium concentration and the emergence of and emotional states make it of great interest to examine thirst in 10 healthy adults. The correlation of regional cerebral whether the cerebellum has a role in basic vegetative functions blood flow with subjects’ ratings of thirst showed major activation and the primal emotions thus generated, particularly given the in the vermal central lobule. -

The Cerebellum in Eye Movement Control
Eye (2015) 29, 191–195 & 2015 Macmillan Publishers Limited All rights reserved 0950-222X/15 www.nature.com/eye CAMBRIDGE OPHTHALMOLOGICAL SYMPOSIUM The cerebellum in VR Patel1 and DS Zee2 eye movement control: nystagmus, coordinate frames and disconjugacy This article has been corrected since Advance Online Publication and an erratum is also printed in this issue Abstract cerebellum in binocular control—both to create disconjugacy when it is necessary, and to In this review we discuss several aspects of prevent ocular misalignment when it is eye movement control in which the unnecessary and disruptive. We will also touch cerebellum is thought to have a key role, but on the implications of evolving into frontal-eyed have been relatively ignored. We will focus creatures, with the competing demands on the mechanisms underlying certain forms of binocular, foveal vs retinal, full-field of cerebellar nystagmus, as well as the stabilization of images. Furthermore, we contributions of the cerebellum to binocular suggest that the phylogenetically old vestibular alignment in healthy and diseased states. anlage persists in a rudimentary form within A contemporary review of our understanding our human brains and its vestiges can be provides a basis for directions of further 1Associate Professor of uncovered in neurological disease.1–4 These inquiry to address some of the uncertainties Ophthalmology, USC Eye issues bear on interpretation of pathological regarding the contributions of the cerebellum Institute, Keck School of nystagmus, which depends on the coordinate Medicine, University of to ocular motor control. system in which the nystagmus is couched Southern California, Eye (2015) 29, 191–195; doi:10.1038/eye.2014.271; Los Angeles, CA, USA (foveal: eye frame vs full-field retina: head published online 14 November 2014 frame), and also on the types of ocular 2Professor of Neurology, misalignment seen in cerebellar patients. -

Spinocerebellar Ataxia Type 6 with Positional Vertigo and Acetazolamide Responsive Episodic Ataxia
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.65.4.565 on 1 October 1998. Downloaded from J Neurol Neurosurg Psychiatry 1998;65:565–568 565 SHORT REPORT Spinocerebellar ataxia type 6 with positional vertigo and acetazolamide responsive episodic ataxia Joanna C Jen, Qing Yue, Juliana Karrim, Stanley F Nelson, Robert W Baloh Abstract tions in other parts of CACNA1A. SCA6 has The SCA6 mutation, a small expansion of clinical features that overlap with those of a CAG repeat in a calcium channel gene EA-2. CACNA1A, was identified in three pedi- grees. Point mutations in other parts of Case reports the gene CACNA1A were excluded and The study was approved by our institutional new clinical features of SCA6 reported— review board and informed consent was namely, central positional nystagmus and obtained from all subjects. The pedigrees are episodic ataxia responsive to acetazola- shown in the figure. mide. The three allelic disorders, episodic ataxia type 2, familial hemiplegic mi- PEDIGREE 1 graine, and SCA6, have overlapping clini- This family was previously reported as an cal features. atypical EA-2 linked to chromosome 19p (J Neurol Neurosurg Psychiatry 1998;65:565–568) (family 4).5 The type of mutation was un- known. The proband began having episodes of Keywords: SCA6; spinocerebellar ataxia; hereditary dizziness and ataxia at the age of 42. She also ataxia; calcium channel noted positional vertigo so that she slept propped up with pillows. About a year later, she noticed mild interictal imbalance. Initial exam- Episodic ataxia type 2 (EA-2) is an autosomal ination disclosed a central type of positional dominant disorder characterised by episodes of nystagmus (conjugate downbeat nystagmus ataxia lasting hours to days with interictal nys- that did not fatigue with repeated positioning). -

Roles of the Declive, Folium, and Tuber Cerebellar Vermian Lobules in Sportspeople
JCN Open Access REVIEW pISSN 1738-6586 / eISSN 2005-5013 / J Clin Neurol 2017 Roles of the Declive, Folium, and Tuber Cerebellar Vermian Lobules in Sportspeople In Sung Parka The cerebellum plays vital roles in balance control and motor learning, including in saccadic Nam Joon Leeb c adaptation and coordination. It consists of the vermis and two hemispheres and is anatomically Im Joo Rhyu separated into ten lobules that are designated as I–X. Although neuroimaging and clinical stud- aDepartment of Liberal Arts, ies suggest that functions are compartmentalized within the cerebellum, the function of each Kyungil University, Gyeongsan, Korea cerebellar lobule is not fully understood. Electrophysiological and lesion studies in animals as b Departments of Diagnostic Radiology and well as neuroimaging and lesion studies in humans have revealed that vermian lobules VI and c Anatomy, Korea University VII (declive, folium, and tuber) are critical for controlling postural balance, saccadic eye move- College of Medicine, Seoul, Korea ments, and coordination. In addition, recent structural magnetic resonance imaging studies have revealed that these lobules are larger in elite basketball and short-track speed skaters. Fur- thermore, in female short-track speed skaters, the volume of this region is significantly correlat- ed with static balance. This article reviews the function of vermian lobules VI and VII, focusing on the control of balance, eye movements, and coordination including coordination between the eyes and hands and bimanual coordination. Key Words balance, cerebellum, coordination, saccade. INTRODUCTION The cerebellum, located in the posterior cranial fossa, is a major motor structure of the brain. It controls motor-related functions, such as maintaining balance and posture, and motor learning including coordination of movements through complex regulatory and feedback mechanisms. -

Zonal Organization of the Flocculovestibular Nucleus Projection in the Rabbit: a Combined Axonal Tracing and Acetylcholinesterase Histochemicalstudy
THE JOURNAL OF COMPARATIVE NEUROLOGY 354k51-71 (1995) Zonal Organization of the Flocculovestibular Nucleus Projection in the Rabbit: A Combined Axonal Tracing and Acetylcholinesterase HistochemicalStudy J. TAN, A.H. EPEMA, AND J. VOOGD Department of Anatomy, Erasmus University Rotterdam, 3000 DR Rotterdam, The Netherlands (J.T., J.V.) and Department of Anesthesiology, Academic Hospital, University of Groningen, Groningen, The Netherlands (A.H.E.) ABSTRACT With the use of retrograde transport of horseradish peroxidase we confirmed the observation of Yamamoto and Shimoyama ([ 19771 Neurosci Lett. 5279-283) that Purkinje cells of the rabbit flocculus projecting to the medial vestibular nucleus are located in two discrete zones, FZIl and FZlv, that alternate with two other Purkinje cell zones, FZI and FZIII, projecting to the superior vestibular nucleus. The retrogradely labeled axons of these Purkinje cells collect in four bundles that occupy the corresponding floccular white matter compart- ments, FC1-4,that can be delineated with acetylcholinesterase histochemistry (Tan et al. [ 1995al J. Comp. Neurol., this issue). Anterograde tracing from small injections of wheat germ agglutin-hoseradish peroxidase in single Purkinje cell zones of the flocculus showed that Purkinje cell axons of FZrr travel in FC2to terminate in the medial vestibular nucleus. Purkinje cell axons from FZI and FZIII occupy the FCI and FC3 compartments, respectively, and terminate in the superior vestibular nucleus. Purkinje cell axons from all three compartments pass through the floccular peduncle and dorsal group y. In addition, some fibers from FZI and FZIJ,but not from FZIII, arch through the cerebellar nuclei to join the floccular peduncle more medially. -

Microvascular Anatomy of the Cerebellar Parafloccular Perforating Space
LABORATORY INVESTIGATION J Neurosurg 124:440–449, 2016 Microvascular anatomy of the cerebellar parafloccular perforating space Pablo Sosa, MD,1 Manuel Dujovny, MD,2 Ibe Onyekachi, BS,2 Noressia Sockwell, BS,2 Fabián Cremaschi, MD,1 and Luis E. Savastano, MD3 1Department of Neuroscience, Clinical and Surgical Neurology, School of Medicine, National University of Cuyo, Mendoza, Argentina; 2Departments of Neurosurgery and Electrical Engineering, Wayne State University, Detroit; and 3Department of Neurosurgery, University of Michigan, Ann Arbor, Michigan OBJECTIVE The cerebellopontine angle is a common site for tumor growth and vascular pathologies requiring surgical manipulations that jeopardize cranial nerve integrity and cerebellar and brainstem perfusion. To date, a detailed study of vessels perforating the cisternal surface of the middle cerebellar peduncle—namely, the paraflocculus or parafloccular perforating space—has yet to be published. In this report, the perforating vessels of the anterior inferior cerebellar artery (AICA) in the parafloccular space, or on the cisternal surface of the middle cerebellar peduncle, are described to eluci- date their relevance pertaining to microsurgery and the different pathologies that occur at the cerebellopontine angle. METHODS Fourteen cadaveric cerebellopontine cisterns (CPCs) were studied. Anatomical dissections and analysis of the perforating arteries of the AICA and posterior inferior cerebellar artery at the parafloccular space were recorded using direct visualization by surgical microscope, optical histology, and scanning electron microscope. A comprehensive review of the English-language and Spanish-language literature was also performed, and findings related to anatomy, histology, physiology, neurology, neuroradiology, microsurgery, and endovascular surgery pertaining to the cerebellar flocculus or parafloccular spaces are summarized. RESULTS A total of 298 perforating arteries were found in the dissected specimens, with a minimum of 15 to a maxi- mum of 26 vessels per parafloccular perforating space.