Description of Brachypelma Annitha N. Sp. and Brachypelma Hamorii N
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sustentable De Especies De Tarántula
Plan de acción de América del Norte para un comercio sustentable de especies de tarántula Comisión para la Cooperación Ambiental Citar como: CCA (2017), Plan de acción de América del Norte para un comercio sustentable de especies de tarántula, Comisión para la Cooperación Ambiental, Montreal, 48 pp. La presente publicación fue elaborada por Rick C. West y Ernest W. T. Cooper, de E. Cooper Environmental Consulting, para el Secretariado de la Comisión para la Cooperación Ambiental. La información que contiene es responsabilidad de los autores y no necesariamente refleja los puntos de vista de los gobiernos de Canadá, Estados Unidos o México. Se permite la reproducción de este material sin previa autorización, siempre y cuando se haga con absoluta precisión, su uso no tenga fines comerciales y se cite debidamente la fuente, con el correspondiente crédito a la Comisión para la Cooperación Ambiental. La CCA apreciará que se le envíe una copia de toda publicación o material que utilice este trabajo como fuente. A menos que se indique lo contrario, el presente documento está protegido mediante licencia de tipo “Reconocimiento – No comercial – Sin obra derivada”, de Creative Commons. Detalles de la publicación Categoría del documento: publicación de proyecto Fecha de publicación: mayo de 2017 Idioma original: inglés Procedimientos de revisión y aseguramiento de la calidad: Revisión final de las Partes: abril de 2017 QA311 Proyecto: Fortalecimiento de la conservación y el aprovechamiento sustentable de especies listadas en el Apéndice II de la -

Lab 8: Arthropoda OEB 51 Lab 8: Arthropoda
Lab 8: Arthropoda OEB 51 Lab 8: Arthropoda 6 April 2016 ATTENTION: Today we will be examining several preserved specimens from along the arthropod diversity and some exciting live specimens that you have not seen in the fieldtrip! Please keep all preserved specimens submerged. Examine using forceps and probes – be careful not to damage or remove the limbs of any specimen. And remember to return the specimen to its original spot in the lateral bench so your friends can find the animals properly labeled. • For each specimen, even when specific instructions are not given, make detailed observations and drawings and take notes on important characters in the evolution of arthropods, such as tagmata and adaptations to the specific environment where the organism lives. 1 Lab 8: Arthropoda OEB 51 Myriapoda With these living myriapods, take the opportunity to make a few notes on behavior – locomotory, hygienic (they frequently clean their antennae). CHOOSE ONE • Chilopoda (live) Do not handle these centipedes directly, as they are venomous (not deadly, but still). Watch these animals move, but don’t let them escape! Try to draw if you can. Compare to the large, preserved Scolopendra gigantea (MCZ specimen). Draw from the preserved for more morphological details. • Diplopoda (live) Contrary to centipedes, millipedes are docile (but they have deterrent substances, like cyanide compounds, which you might be able to smell after handling the specimens). Observe their movement. Make a sketch of the whole body and drawings of specific parts in more detail. • What is the most conspicuous difference between these two groups of myriapods? 2 Lab 8: Arthropoda OEB 51 Pycnogonida • Colossendeis colossea (Museum specimens, handle with care) • What characteristics of pycnogonids are not found in other arthropods? 3 Lab 8: Arthropoda OEB 51 Chelicerata • Xiphosura – Limulus (horseshoe crab). -

BMB-WRC Animal Inventory
Department of Environment and Natural Resources BIODIVERSITY MANAGEMENT BUREAU Quezon Avenue, Diliman, Quezon City INVENTORY OF LIVE ANIMALS AT THE BMB-WILDLIFE RESCUE CENTER AS OF JULY 31, 2020 SPECIES STOCK ON HAND (AS OF COMMON NAME SCIENTIFIC NAME JULY 31, 2020) MAMMALS ENDEMIC / INDIGENOUS 1. Northern luzon cloud Ploeomys pallidus 1 rat 2. Palawan bearcat Arctictis binturong 2 3. Philippine deer Rusa marianna 2 4. Philippine monkey or Macaca fascicularis 92 Long-tailed macaque 5. Philippine palm civet Paradoxurus hermaphroditus 6 EXOTIC 6. Hedgehog Atelerix frontalis 1 7. Serval cat Leptailurus serval 2 8. Sugar glider Petaurus breviceps 58 9. Tiger Panthera tigris 2 10. Vervet monkey Chlorocebus pygerythrus 1 11. White handed gibbon Hylobates lar 1 Sub-total A 168 (Mammals) AVIANS ENDEMIC / INDIGENOUS 12. Black kite Milvus migrans 1 13. Black-crowned night Nycticorax nycticorax 1 heron 14. Blue-naped parrot Tanygnathus lucionensis 4 15. Brahminy kite Haliastur indus 41 16. Changeable hawk Spizaetus cirrhatus 6 eagle 17. Crested goshawk Accipiter trivirgatus 1 18. Crested serpent eagle Spilornis cheela 24 19. Green imperial pigeon Ducula aenea 2 20. Grey-headed fish eagle Haliaeetus ichthyaetus 1 21. Nicobar pigeon Caloenas nicobarica 1 22. Palawan hornbill Anthracoceros marchei 2 23. Palawan talking myna Gracula religiosa 3 24. Philippine eagle Pithecophaga jefferyi 1 25. Philippine hanging Loriculus philippensis 11 parrot 26. Philippine hawk eagle Spizaetus philippensis 12 27. Philippine horned Bubo philippensis 9 (eagle) owl 28. Philippine Scops owl Otus megalotis 5 29. Pink-necked pigeon Treron vernans 1 30. Pinsker's hawk eagle Spizaetus pinskerii 1 31. Red turtle dove Streptopelia tranquebarica 1 32. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -

Tarantulas and Social Spiders
Tarantulas and Social Spiders: A Tale of Sex and Silk by Jonathan Bull BSc (Hons) MSc ICL Thesis Presented to the Institute of Biology of The University of Nottingham in Partial Fulfilment of the Requirements for the Degree of Doctor of Philosophy The University of Nottingham May 2012 DEDICATION To my parents… …because they both said to dedicate it to the other… I dedicate it to both ii ACKNOWLEDGEMENTS First and foremost I would like to thank my supervisor Dr Sara Goodacre for her guidance and support. I am also hugely endebted to Dr Keith Spriggs who became my mentor in the field of RNA and without whom my understanding of the field would have been but a fraction of what it is now. Particular thanks go to Professor John Brookfield, an expert in the field of biological statistics and data retrieval. Likewise with Dr Susan Liddell for her proteomics assistance, a truly remarkable individual on par with Professor Brookfield in being able to simplify even the most complex techniques and analyses. Finally, I would really like to thank Janet Beccaloni for her time and resources at the Natural History Museum, London, permitting me access to the collections therein; ten years on and still a delight. Finally, amongst the greats, Alexander ‘Sasha’ Kondrashov… a true inspiration. I would also like to express my gratitude to those who, although may not have directly contributed, should not be forgotten due to their continued assistance and considerate nature: Dr Chris Wade (five straight hours of help was not uncommon!), Sue Buxton (direct to my bench creepy crawlies), Sheila Keeble (ventures and cleans where others dare not), Alice Young (read/checked my thesis and overcame her arachnophobia!) and all those in the Centre for Biomolecular Sciences. -
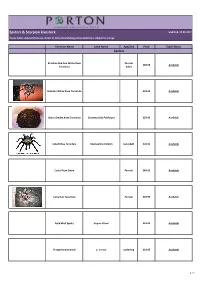
Spiders & Scorpion Livestock
Spiders & Scorpion Livestock Updated: 22.09.2017 Please Note: Livestock lists are correct at time of publishing and availability is subject to change Common Name Latin Name Age/Size Price Stock Status Spiders Brazilian Red And White Knee Female £49.95 Available Tarantula Adult Brazilian White Knee Tarantula £34.95 Available Chaco Golden Knee Tarantula Grammostola Pulchripes £29.95 Available Cobalt Blue Tarantula Haplopelma Lividum Sub Adult £49.95 Available Costa Rican Zebra Female £49.95 Available Curly Hair Tarantula Female £29.95 Available Field Wolf Spider Hogna Miami £19.95 Available Fringed Ornamental p. ornata spiderling £24.95 Available 1 / 4 Common Name Latin Name Age/Size Price Stock Status Spiders Golden Baboon Tarantula Augacephalus ezendami £39.95 Available Gooty Ornamental p. metallica spiderling £49.95 Available Green Bottle Blue Tarantula Female £49.95 Available Green Femur Birdeater Phormictopus"Green Femur" £39.95 Available Indian Ornamental Tarantula Poecilotheria regalis 2cm £24.95 Available King Baboon Tarantula Pelinobius Muticus £24.95 Available Martinique Pink Toe Tarantula 2cm £16.95 Available Mexican Fire Leg Tarantula Brachypelma Bohemi £39.95 Available Mexican Red Knee Tarantula Brachypelma Smithi £34.95 Available 2 / 4 Common Name Latin Name Age/Size Price Stock Status Spiders Mexican Red Leg Tarantula Brachypelma Emilia £39.95 Available £24.95 Available Mexican Red Rump Brachypelma Vagans Female Sub Adult Available £49.95 spiderling £19.95 Available OBT Tarantula Female Sub Adult Available £39.95 Purple Earth -

Genetic Analysis of Tarantulas in the Genus Brachypelma Using Inter Simple Sequence Repeats (ISSR)
Eastern Michigan University DigitalCommons@EMU Senior Honors Theses & Projects Honors College 2020 Genetic analysis of tarantulas in the genus Brachypelma using Inter Simple Sequence Repeats (ISSR) Sarah Holtzen Follow this and additional works at: https://commons.emich.edu/honors Part of the Biology Commons Recommended Citation Holtzen, Sarah, "Genetic analysis of tarantulas in the genus Brachypelma using Inter Simple Sequence Repeats (ISSR)" (2020). Senior Honors Theses & Projects. 688. https://commons.emich.edu/honors/688 This Open Access Senior Honors Thesis is brought to you for free and open access by the Honors College at DigitalCommons@EMU. It has been accepted for inclusion in Senior Honors Theses & Projects by an authorized administrator of DigitalCommons@EMU. For more information, please contact [email protected]. Genetic analysis of tarantulas in the genus Brachypelma using Inter Simple Sequence Repeats (ISSR) Abstract There is a great deal of morphological and genetic species diversity on Earth that requires careful conservation. One such genetically diverse genus of tarantulas is that of Brachypelma. In this study, we employ a newer DNA fingerprinting technique known as Inter Simple Sequence Repeat (ISSR), ot study the genetic variation among Brachypelma species and to determine if the invasive Brachypelma tarantula found in Florida B. vagans. Although B. vagans is a species protected under CITES Appendix II, this species has a wide distribution in Mexico and traits allowing for invasion to new habitats. It was hypothesized that the invasive tarantula in Florida is that of B. vagans and that it would be more closely related to samples from the Mexican populations as opposed to samples from the United States pet trade. -

Husbandry Manual for Exotic Tarantulas
Husbandry Manual for Exotic Tarantulas Order: Araneae Family: Theraphosidae Author: Nathan Psaila Date: 13 October 2005 Sydney Institute of TAFE, Ultimo Course: Zookeeping Cert. III 5867 Lecturer: Graeme Phipps Table of Contents Introduction 6 1 Taxonomy 7 1.1 Nomenclature 7 1.2 Common Names 7 2 Natural History 9 2.1 Basic Anatomy 10 2.2 Mass & Basic Body Measurements 14 2.3 Sexual Dimorphism 15 2.4 Distribution & Habitat 16 2.5 Conservation Status 17 2.6 Diet in the Wild 17 2.7 Longevity 18 3 Housing Requirements 20 3.1 Exhibit/Holding Area Design 20 3.2 Enclosure Design 21 3.3 Spatial Requirements 22 3.4 Temperature Requirements 22 3.4.1 Temperature Problems 23 3.5 Humidity Requirements 24 3.5.1 Humidity Problems 27 3.6 Substrate 29 3.7 Enclosure Furnishings 30 3.8 Lighting 31 4 General Husbandry 32 4.1 Hygiene and Cleaning 32 4.1.1 Cleaning Procedures 33 2 4.2 Record Keeping 35 4.3 Methods of Identification 35 4.4 Routine Data Collection 36 5 Feeding Requirements 37 5.1 Captive Diet 37 5.2 Supplements 38 5.3 Presentation of Food 38 6 Handling and Transport 41 6.1 Timing of Capture and handling 41 6.2 Catching Equipment 41 6.3 Capture and Restraint Techniques 41 6.4 Weighing and Examination 44 6.5 Transport Requirements 44 6.5.1 Box Design 44 6.5.2 Furnishings 44 6.5.3 Water and Food 45 6.5.4 Release from Box 45 7 Health Requirements 46 7.1 Daily Health Checks 46 7.2 Detailed Physical Examination 47 7.3 Chemical Restraint 47 7.4 Routine Treatments 48 7.5 Known Health Problems 48 7.5.1 Dehydration 48 7.5.2 Punctures and Lesions 48 7.5.3 -

Fall 2009 3 Academy Abbreviated a Bite of Evolution Locking Teeth
greetings from the academy In the pages of Academy Fron- tions are challenged financially, and some have been unable to tiers, we hope you give you an meet that challenge. in-depth look inside the museum, particularly into our treasured col- A grant from the Andrew Mellon Foundation funded an inter- lections, the lifeblood of the Acad- national gathering this past June at the Academy of the chief emy. The stories emerging from executives of eight leading natural history institutions located our collections are endless—from in the United States, United Kingdom, Paris, the Netherlands, invaluable specimens collected by and South Africa. Lewis and Clark on their 3,700- mile expedition, to the recent dis- We discussed possible new markets for science collections and Bruce Tepper/ ANSP covery of a new species stored in we identified as top priorities the need to preserve viable tissue the shadows of the collection for more than 150 years. and DNA of all known species, image and digitize the infor- mation for all type specimens, and generally improve coordi- These collections find many uses. Their study is the backbone nation and access to museum information. of research in systematics and evolutionary biology. They are used by conservation and environmental protection agencies to This meeting is only the beginning—it’s the first of what we document and monitor success in forestry management, endan- hope will be a continued collaboration in this effort to share gered species protection, and invasive species prevention and our collections with the world. Stay tuned! control. They provide the basis for understanding the diversity of life, and the identification of new species worldwide. -

Spider Notebooking Pages Bw
Spider Notebooking Pages 2019 Stacey Jones at Simple Living. Creative Learning PINTEREST! | WEBSITE | FACEBOOK All rights reserved. No part of this book may be reproduced, stored or transmitted in any form by any means without prior permission of the publisher. This workbook is licensed for personal/family use only. YOU MAY: ! Use these files for personal use only. ! Use in your personal classroom ! Download the files to your personal computer. ! Print as many copies as you would like to use for your personal use. ! Direct other to our website: https://simplelivingcreativelearning.com YOU MAY NOT: ! Edit any of these printables. ! Share the files with anyone else. ! Store or sell them on any website. ! Claim them as your own. ! Print and sell or distribute them to others Graphics and Font: Spider Notebooking Pages Spider Notebooking Pages Spider Notebooking Pages By: !!!!!!!!!!!!!!!!!!!!!!!! Spider Notebooking Pages By: !!!!!!!!!!!!!!!!!!!!!!!! Spider Notebooking Pages By: !!!!!!!!!!!!!!!!!!!!!!!! Arrow-shaped micrathena (Micrathena sagittata) Diet Habitat Characteristics Simple Living. Creative Learning ! Arrow-shaped micrathena (Micrathena sagittata) ______________ ________0__000 00000000000000 Scientific Classification Order: ______________ ________________________ Suborder: ____________ Genus: _______________ ________________________ Species: _____________ ________________________ ________________________ ________________________ ________________________ ________________________ ________________________ ________________________ ________________________ -
Chamberlin & Ivie, 1936
A peer-reviewed open-access journal ZooKeys 526:Revised 75–104 generic(2015) placement of Brachypelma embrithes (Chamberlin & Ivie, 1936)... 75 doi: 10.3897/zookeys.526.6315 RESEARCH ARTICLE http://zookeys.pensoft.net Launched to accelerate biodiversity research Revised generic placement of Brachypelma embrithes (Chamberlin & Ivie, 1936) and Brachypelma angustum Valerio, 1980, with definition of the taxonomic features for identification of female Sericopelma Ausserer, 1875 (Araneae, Theraphosidae) Ray Gabriel1, Stuart J. Longhorn1 1 Hope Entomological Collections, Oxford University Museum of Natural History (OUMNH), Parks Road, Oxford, England, OX1 3PW, United Kingdom Corresponding author: Stuart J. Longhorn ([email protected]) Academic editor: J. Miller | Received 24 August 2015 | Accepted 22 September 2015 | Published 12 October 2015 http://zoobank.org/BA29348F-1339-413E-8F16-B08F78DB167D Citation: Gabriel R, Longhorn SJ (2015) Revised generic placement of Brachypelma embrithes (Chamberlin & Ivie, 1936) and Brachypelma angustum Valerio, 1980, with definition of the taxonomic features for identification of femaleSericopelma Ausserer, 1875 (Araneae, Theraphosidae). ZooKeys 526: 75–104. doi: 10.3897/zookeys.526.6315 Abstract The tarantula genusSericopelma was originally defined based on male specimens, most notably lacking tibial spurs on leg I. Early female specimens were unrecognised as Sericopelma, and typically placed in Eurypelma – a dumping ground for problem specimens. The first females were only later recognised, but authors failed to adequately define femaleSericopelma . Here, the holotypes of the Southern-most alleged Brachypelma species, B. embrithes (Chamberlin & Ivie, 1936) and B. angustum Valerio, 1980 were exam- ined, and finding both to possess defining characteristics of Sericopelma were transferred. The taxonomic attributes to define Sericopelma relative to Brachypelma and select other Neotropical genera are discussed, especially for females. -

Morphology, Evolution and Usage of Urticating Setae by Tarantulas (Araneae: Theraphosidae)
ZOOLOGIA 30 (4): 403–418, August, 2013 http://dx.doi.org/10.1590/S1984-46702013000400006 Morphology, evolution and usage of urticating setae by tarantulas (Araneae: Theraphosidae) Rogério Bertani1,3 & José Paulo Leite Guadanucci2 1 Laboratório Especial de Ecologia e Evolução, Instituto Butantan. Avenida Vital Brazil 1500, 05503-900 São Paulo SP, Brazil. E-mail: [email protected] 2 Laboratório de Zoologia de Invertebrados, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Campus JK. Rodovia MGT 367, km 583, 39100-000 Diamantina, MG, Brazil. E-mail: [email protected] 3 Corresponding author. ABSTRACT. Urticating setae are exclusive to New World tarantulas and are found in approximately 90% of the New World species. Six morphological types have been proposed and, in several species, two morphological types can be found in the same individual. In the past few years, there has been growing concern to learn more about urticating setae, but many questions still remain unanswered. After studying individuals from several theraphosid species, we endeavored to find more about the segregation of the different types of setae into different abdominal regions, and the possible existence of patterns; the morphological variability of urticating setae types and their limits; whether there is variability in the length of urticating setae across the abdominal area; and whether spiders use different types of urticat- ing setae differently. We found that the two types of urticating setae, which can be found together in most theraphosine species, are segregated into distinct areas on the spider’s abdomen: type III occurs on the median and posterior areas with either type I or IV surrounding the patch of type III setae.