Incorporating the Microbial Loop Simple Plankton Model 1N A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Response of Marine Food Webs to Climate-Induced Changes in Temperature and Inflow of Allochthonous Organic Matter
Response of marine food webs to climate-induced changes in temperature and inflow of allochthonous organic matter Rickard Degerman Department of Ecology and Environmental Science 901 87 Umeå Umeå 2015 1 Copyright©Rickard Degerman ISBN: 978-91-7601-266-6 Front cover illustration by Mats Minnhagen Printed by: KBC Service Center, Umeå University Umeå, Sweden 2015 2 Tillägnad Maria, Emma och Isak 3 Table of Contents Abstract 5 List of papers 6 Introduction 7 Aquatic food webs – different pathways Food web efficiency – a measure of ecosystem function Top predators cause cascade effects on lower trophic levels The Baltic Sea – a semi-enclosed sea exposed to multiple stressors Varying food web structures Climate-induced changes in the marine ecosystem Food web responses to increased temperature Responses to inputs of allochthonous organic matter Objectives 14 Material and Methods 14 Paper I Paper II and III Paper IV Results and Discussion 18 Effect of temperature and nutrient availability on heterotrophic bacteria Influence of food web length and labile DOC on pelagic productivity and FWE Consequences of changes in inputs of ADOM and temperature for pelagic productivity and FWE Control of pelagic productivity, FWE and ecosystem trophic balance by colored DOC Conclusion and future perspectives 21 Author contributions 23 Acknowledgements 23 Thanks 24 References 25 4 Abstract Global records of temperature show a warming trend both in the atmosphere and in the oceans. Current climate change scenarios indicate that global temperature will continue to increase in the future. The effects will however be very different in different geographic regions. In northern Europe precipitation is projected to increase along with temperature. -

7.014 Handout PRODUCTIVITY: the “METABOLISM” of ECOSYSTEMS
7.014 Handout PRODUCTIVITY: THE “METABOLISM” OF ECOSYSTEMS Ecologists use the term “productivity” to refer to the process through which an assemblage of organisms (e.g. a trophic level or ecosystem assimilates carbon. Primary producers (autotrophs) do this through photosynthesis; Secondary producers (heterotrophs) do it through the assimilation of the organic carbon in their food. Remember that all organic carbon in the food web is ultimately derived from primary production. DEFINITIONS Primary Productivity: Rate of conversion of CO2 to organic carbon (photosynthesis) per unit surface area of the earth, expressed either in terns of weight of carbon, or the equivalent calories e.g., g C m-2 year-1 Kcal m-2 year-1 Primary Production: Same as primary productivity, but usually expressed for a whole ecosystem e.g., tons year-1 for a lake, cornfield, forest, etc. NET vs. GROSS: For plants: Some of the organic carbon generated in plants through photosynthesis (using solar energy) is oxidized back to CO2 (releasing energy) through the respiration of the plants – RA. Gross Primary Production: (GPP) = Total amount of CO2 reduced to organic carbon by the plants per unit time Autotrophic Respiration: (RA) = Total amount of organic carbon that is respired (oxidized to CO2) by plants per unit time Net Primary Production (NPP) = GPP – RA The amount of organic carbon produced by plants that is not consumed by their own respiration. It is the increase in the plant biomass in the absence of herbivores. For an entire ecosystem: Some of the NPP of the plants is consumed (and respired) by herbivores and decomposers and oxidized back to CO2 (RH). -

Bacterial Production and Respiration
Organic matter production % 0 Dissolved Particulate 5 > Organic Organic Matter Matter Heterotrophic Bacterial Grazing Growth ~1-10% of net organic DOM does not matter What happens to the 90-99% of sink, but can be production is physically exported to organic matter production that does deep sea not get exported as particles? transported Export •Labile DOC turnover over time scales of hours to days. •Semi-labile DOC turnover on time scales of weeks to months. •Refractory DOC cycles over on time scales ranging from decadal to multi- decadal…perhaps longer •So what consumes labile and semi-labile DOC? How much carbon passes through the microbial loop? Phytoplankton Heterotrophic bacteria ?? Dissolved organic Herbivores ?? matter Higher trophic levels Protozoa (zooplankton, fish, etc.) ?? • Very difficult to directly measure the flux of carbon from primary producers into the microbial loop. – The microbial loop is mostly run on labile (recently produced organic matter) - - very low concentrations (nM) turning over rapidly against a high background pool (µM). – Unclear exactly which types of organic compounds support bacterial growth. Bacterial Production •Step 1: Determine how much carbon is consumed by bacteria for production of new biomass. •Bacterial production (BP) is the rate that bacterial biomass is created. It represents the amount of Heterotrophic material that is transformed from a nonliving pool bacteria (DOC) to a living pool (bacterial biomass). •Mathematically P = µB ?? µ = specific growth rate (time-1) B = bacterial biomass (mg C L-1) P= bacterial production (mg C L-1 d-1) Dissolved organic •Note that µ = P/B matter •Thus, P has units of mg C L-1 d-1 Bacterial production provides one measurement of carbon flow into the microbial loop How doe we measure bacterial production? Production (∆ biomass/time) (mg C L-1 d-1) • 3H-thymidine • 3H or 14C-leucine Note: these are NOT direct measures of biomass production (i.e. -
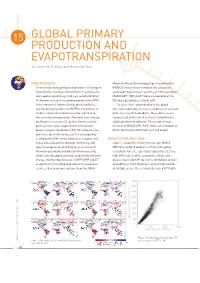
GLOBAL PRIMARY PRODUCTION and EVAPOTRANSPIRATION by Steven W
15 GLOBAL PRIMARY PRODUCTION AND EVAPOTRANSPIRATION by Steven W. Running and Maosheng Zhao DATA PRODUCTS Moderate Resolution Imaging Spectroradiometer Terrestrial primary production provides the energy to (MODIS) sensor, these variables are calculated maintain the structure and functions of ecosystems, every eight days in near real time at 1 km resolution. and supplies goods (e.g. food, fuel, wood and fi bre) MODIS GPP, NPP and ET data are available at the for human society. Gross primary production (GPP) EOS data gateway (see link below). is the amount of carbon fi xed by photosynthesis, To correct for contamination in the global and net primary production (NPP) is the amount of refl ectance data due to severe cloudiness or aerosols carbon converted into biomass after subtracting in the near real time products, these datasets are the cost of plant respiration. The water loss through reprocessed at the end of each year to build more exchange of trace gas CO2 by leaf stomata during stable, permanent datasets. These end-of-year photosynthesis plus evaporation from soil and versions of MODIS GPP, NPP and ET are available at plants is evapotranspiration (ET). ET computes the NTSG, University of Montana (see link below). water lost by a land surface, so it is consequently a component of the water balance in a region, and RESULTS FOR 2000–2006 is therefore relevant for drought monitoring and Figure 1 shows the seven-year average MODIS water management, providing an assessment of NPP for vegetated land on earth at 1 km spatial the water potentially available for human society. -

Phytoplankton As Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate
sustainability Review Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate Samarpita Basu * ID and Katherine R. M. Mackey Earth System Science, University of California Irvine, Irvine, CA 92697, USA; [email protected] * Correspondence: [email protected] Received: 7 January 2018; Accepted: 12 March 2018; Published: 19 March 2018 Abstract: The world’s oceans are a major sink for atmospheric carbon dioxide (CO2). The biological carbon pump plays a vital role in the net transfer of CO2 from the atmosphere to the oceans and then to the sediments, subsequently maintaining atmospheric CO2 at significantly lower levels than would be the case if it did not exist. The efficiency of the biological pump is a function of phytoplankton physiology and community structure, which are in turn governed by the physical and chemical conditions of the ocean. However, only a few studies have focused on the importance of phytoplankton community structure to the biological pump. Because global change is expected to influence carbon and nutrient availability, temperature and light (via stratification), an improved understanding of how phytoplankton community size structure will respond in the future is required to gain insight into the biological pump and the ability of the ocean to act as a long-term sink for atmospheric CO2. This review article aims to explore the potential impacts of predicted changes in global temperature and the carbonate system on phytoplankton cell size, species and elemental composition, so as to shed light on the ability of the biological pump to sequester carbon in the future ocean. -

Microbial Loop' in Stratified Systems
MARINE ECOLOGY PROGRESS SERIES Vol. 59: 1-17, 1990 Published January 11 Mar. Ecol. Prog. Ser. 1 A steady-state analysis of the 'microbial loop' in stratified systems Arnold H. Taylor, Ian Joint Plymouth Marine Laboratory, Prospect Place, West Hoe, Plymouth PLl 3DH, United Kingdom ABSTRACT. Steady state solutions are presented for a simple model of the surface mixed layer, which contains the components of the 'microbial loop', namely phytoplankton, picophytoplankton, bacterio- plankton, microzooplankton, dissolved organic carbon, detritus, nitrate and ammonia. This system is assumed to be in equilibrium with the larger grazers present at any time, which are represented as an external mortality function. The model also allows for dissolved organic nitrogen consumption by bacteria, and self-grazing and mixotrophy of the microzooplankton. The model steady states are always stable. The solution shows a number of general properties; for example, biomass of each individual component depends only on total nitrogen concentration below the mixed layer, not whether the nitrogen is in the form of nitrate or ammonia. Standing stocks and production rates from the model are compared with summer observations from the Celtic Sea and Porcupine Sea Bight. The agreement is good and suggests that the system is often not far from equilibrium. A sensitivity analysis of the model is included. The effect of varying the mixing across the pycnocline is investigated; more intense mixing results in the large phytoplankton population increasing at the expense of picophytoplankton, micro- zooplankton and DOC. The change from phytoplankton to picophytoplankton dominance at low mixing occurs even though the same physiological parameters are used for both size fractions. -

Developments in Aquatic Microbiology
INTERNATL MICROBIOL (2000) 3: 203–211 203 © Springer-Verlag Ibérica 2000 REVIEW ARTICLE Samuel P. Meyers Developments in aquatic Department of Oceanography and Coastal Sciences, Louisiana State University, microbiology Baton Rouge, LA, USA Received 30 August 2000 Accepted 29 September 2000 Summary Major discoveries in marine microbiology over the past 4-5 decades have resulted in the recognition of bacteria as a major biomass component of marine food webs. Such discoveries include chemosynthetic activities in deep-ocean ecosystems, survival processes in oligotrophic waters, and the role of microorganisms in food webs coupled with symbiotic relationships and energy flow. Many discoveries can be attributed to innovative methodologies, including radioisotopes, immunofluores- cent-epifluorescent analysis, and flow cytometry. The latter has shown the key role of marine viruses in marine system energetics. Studies of the components of the “microbial loop” have shown the significance of various phagotrophic processes involved in grazing by microinvertebrates. Microbial activities and dissolved organic carbon are closely coupled with the dynamics of fluctuating water masses. New biotechnological approaches and the use of molecular biology techniques still provide new and relevant information on the role of microorganisms in oceanic and estuarine environments. International interdisciplinary studies have explored ecological aspects of marine microorganisms and their significance in biocomplexity. Studies on the Correspondence to: origins of both life and ecosystems now focus on microbiological processes in the Louisiana State University Station. marine environment. This paper describes earlier and recent discoveries in marine Post Office Box 19090-A. Baton Rouge, LA 70893. USA (aquatic) microbiology and the trends for future work, emphasizing improvements Tel.: +1-225-3885180 in methodology as major catalysts for the progress of this broadly-based field. -

Phytoplankton Primary Production and Its Utilization by the Pelagic Community in the Coastal Zone of the Gulf of Gdahsk (Southern Baltic)
MARINE ECOLOGY PROGRESS SERIES Vol. 148: 169-186. 1997 Published March 20 Mar Ecol Prog Ser l Phytoplankton primary production and its utilization by the pelagic community in the coastal zone of the Gulf of Gdahsk (southern Baltic) Zbigniew Witekl-*,Stanislaw Ochockil, Modesta ~aciejowska~, Marianna Pastuszakl, Jan ~akonieczny',Beata podgorska2, Janina M. ~ownacka', Tomasz Mackiewiczl, Magdalena Wrzesinska-Kwiecieh' 'Sea Fisheries Institute, ul. Koltqtaja 1, 81-332 Gdynia, Poland 'Marine Biology Center, Polish Academy of Sciences, ul. SW. Wojciecha S,81-347 Gdynia, Poland ABSTRACT In th~sstudy we estimated the amount and fate of phytoplankton primary product~onin the coastal zone of the Gulf of Gdansk, Poland, an area exposed to nutnent enrichment from the Vis- tuld R~verand nearby inunlcipal agglomeration The ~nvestigat~onswere carned out at 2 sites dur~ng 5 months in 1993 (February, April, May, August and October). A prolonged bloom pei-iod occurred in the coastal zone, as compared to the open Gulf and the open sea waters. From Aprll until October most values of gross primary production in the near-surface layer were in the range 100 to 500 mgC m-'' d1 Phytoplankton net exudate release constituted on average 5% of the gross prlmai-y production, total exudate release was estimated to be about 2 times h~gher.Bacterial production in the growth season was relatively low (the mean value ly~ngbetween 5 and 9%of gloss primary production), nevertheless, the microbial community (bactena and protozoans) ut~l~zeda large proportion of primary production (from about 50% in April and May to 16'%, in October). -

Controls and Structure of the Microbial Loop
Controls and Structure of the Microbial Loop A symposium organized by the Microbial Oceanography summer course sponsored by the Agouron Foundation Saturday, July 1, 2006 Asia Room, East-West Center, University of Hawaii Symposium Speakers: Peter J. leB Williams (University of Bangor, Wales) David L. Kirchman (University of Delaware) Daniel J. Repeta (Woods Hole Oceanographic Institute) Grieg Steward (University of Hawaii) The oceans constitute the largest ecosystems on the planet, comprising more than 70% of the surface area and nearly 99% of the livable space on Earth. Life in the oceans is dominated by microbes; these small, singled-celled organisms constitute the base of the marine food web and catalyze the transformation of energy and matter in the sea. The microbial loop describes the dynamics of microbial food webs, with bacteria consuming non-living organic matter and converting this energy and matter into living biomass. Consumption of bacteria by predation recycles organic matter back into the marine food web. The speakers of this symposium will explore the processes that control the structure and functioning of microbial food webs and address some of these fundamental questions: What aspects of microbial activity do we need to measure to constrain energy and material flow into and out of the microbial loop? Are we able to measure bacterioplankton dynamics (biomass, growth, production, respiration) well enough to edu/agouroninstitutecourse understand the contribution of the microbial loop to marine systems? What factors control the flow of material and energy into and out of the microbial loop? At what scales (space and time) do we need to measure processes controlling the growth and metabolism of microorganisms? How does our knowledge of microbial community structure and diversity influence our understanding of the function of the microbial loop? Program: 9:00 am Welcome and Introductory Remarks followed by: Peter J. -

Sinking Jelly-Carbon Unveils Potential Environmental Variability Along a Continental Margin
Sinking Jelly-Carbon Unveils Potential Environmental Variability along a Continental Margin Mario Lebrato1,2*, Juan-Carlos Molinero1, Joan E. Cartes3, Domingo Lloris3, Fre´de´ric Me´lin4, Laia Beni- Casadella3 1 Department of Biogeochemistry and Ecology, Helmholtz Centre for Ocean Research Kiel (GEOMAR), Kiel, Germany, 2 Department of Geosciences, Scripps Institution of Oceanography, San Diego, California, United States of America, 3 Institut de Cie`ncies del Mar de Barcelona (CSIC), Barcelona, Spain, 4 Joint Research Centre, Ispra, Italy Abstract Particulate matter export fuels benthic ecosystems in continental margins and the deep sea, removing carbon from the upper ocean. Gelatinous zooplankton biomass provides a fast carbon vector that has been poorly studied. Observational data of a large-scale benthic trawling survey from 1994 to 2005 provided a unique opportunity to quantify jelly-carbon along an entire continental margin in the Mediterranean Sea and to assess potential links with biological and physical variables. Biomass depositions were sampled in shelves, slopes and canyons with peaks above 1000 carcasses per trawl, translating to standing stock values between 0.3 and 1.4 mg C m2 after trawling and integrating between 30,000 and 175,000 m2 of seabed. The benthopelagic jelly-carbon spatial distribution from the shelf to the canyons may be explained by atmospheric forcing related with NAO events and dense shelf water cascading, which are both known from the open Mediterranean. Over the decadal scale, we show that the jelly-carbon depositions temporal variability paralleled hydroclimate modifications, and that the enhanced jelly-carbon deposits are connected to a temperature-driven system where chlorophyll plays a minor role. -

01 Delong 2-8 6/9/05 8:58 AM Page 2
01 Delong 2-8 6/9/05 8:58 AM Page 2 INSIGHT REVIEW NATURE|Vol 437|15 September 2005|doi:10.1038/nature04157 Genomic perspectives in microbial oceanography Edward F. DeLong1 and David M. Karl2 The global ocean is an integrated living system where energy and matter transformations are governed by interdependent physical, chemical and biotic processes. Although the fundamentals of ocean physics and chemistry are well established, comprehensive approaches to describing and interpreting oceanic microbial diversity and processes are only now emerging. In particular, the application of genomics to problems in microbial oceanography is significantly expanding our understanding of marine microbial evolution, metabolism and ecology. Integration of these new genome-enabled insights into the broader framework of ocean science represents one of the great contemporary challenges for microbial oceanographers. Marine ecosystems are complex and dynamic. A mechanistic under- greatly aid in these efforts. The correlation between organism- and habi- standing of the susceptibility of marine ecosystems to global environ- tat-specific genomic features and other physical, chemical and biotic mental variability and climate change driven by greenhouse gases will variables has the potential to refine our understanding of microbial and require a comprehensive description of several factors. These include biogeochemical process in ocean systems. marine physical, chemical and biological interactions including All these advances — improved cultivation, environmental genomic thresholds, negative and positive feedback mechanisms and other approaches and in situ microbial observatories — promise to enhance nonlinear interactions. The fluxes of matter and energy, and the our understanding of the living ocean system. Below, we provide a brief microbes that mediate them, are of central importance in the ocean, recent history of marine microbiology and outline some of the recent yet remain poorly understood. -

Lecture 13 - Primary Production: Water Column Processes Prof
12.742 - Marine Chemistry Fall 2004 Lecture 13 - Primary Production: Water Column Processes Prof. Scott Doney Somewhat different organization from years past - start with surface productivity and work • our way down the water column Try to fold methods in as we go • Electronic notes still missing many of the figures - also somewhat different order. • Primary Production – Formation and respiration of organic matter Photosynthesis / respiration – Formation and dissolution of CaCO3, SiO3 Remineralization – Transport of dust, resuspended sediments, Dom Export particles – Scavenging onto particles Sinking Export – Elemental stoichiometry – Water column distributions – Rates Figure 1. hv Phototrophy – Categorize organisms by source of energy, inorganic electron donor and carbon source. CO + H O C + O 2 2 org 2 Chemo litho autotrophy · Chemo organo heterotrophy · Respiration Photo litho autotrophy Q (Chemical Energy) · Figure 2. 1 N N Light absorbed by pigments. • M – mainly chlorophyll a N N – also accessory pigments (which absorb different wavelengths) including forms of chlorophyll (b, c1, c2) carotenoids, biliopro- teins Figure 3. – Chlorophyll has conjugated double bonds - non-localized π orbital electrons, can absorb solar radiation, bump up to higher electron state – PAR - photosynthetic available radiation (350 700 nm) − – Convert light energy into electron energy 400 λ (nm) 700 – Light harvesting pigment-protein complexes Absorb in Blue and Red (leave Green) involved in electron transfer, funneling ex- cited electrons to reaction sites.