Apple Pollination Biology for Stable and Novel Fruit Production: Search
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Use of Plant Growth Regulators for Branching of Nursery Trees in NY State Mario Miranda Sazo1 and Terence L
The Use of Plant Growth Regulators for Branching of Nursery Trees in NY State Mario Miranda Sazo1 and Terence L. Robinson2 1Cornell Cooperative Extension, Lake Ontario Fruit Program, Newark, NY 2Department of Horticulture, Cornell University, Geneva, NY he quality of nursery trees has a large impact on early pro- the use of Maxcel has been adopted by Italian nurserymen and is a duction and profitability of high density systems. Today, key step for the successful production of well feathered “knip-boom” nurserymen are not only asked to produce trees of good cali- apple trees. Depending on cultivar, Italian nurserymen apply from 1 T per but also highly to 4 Maxcel treatments with spray intervals of 5-7 days depending A new plant growth regulator, cyclanilide branched trees for on temperatures after application (Dr. Walter Guerra, Italy, personal “ the Tall Spindle communication). Since 2009, a new branching agent, cyclanilide (Tradename=Tiberon) very effectively system with short, (Tiberon formulated by Bayer Environmental Science, N.C. USA), induced lateral branching in nursery well positioned lat- has been available in the US for use in outdoor nurseries of apple, trees of 4 apple cultivars in the warm eral branches with sweet cherry, pear, and plum in Florida, Idaho, Oregon, Michigan, and humid year of 2010 in NY State. wide crotch angles. and Washington States. It is not currently registered for tree fruit This has required nursery use in New York State, Europe, or elsewhere. However, it also stopped shoot growth nurseries to im- Studies of the effects of Tiberon for branch induction of nursery for several weeks and reduced final prove their man- apple trees (and other fruit species) were conducted by Dr. -

Apples Catalogue 2019
ADAMS PEARMAIN Herefordshire, England 1862 Oct 15 Nov Mar 14 Adams Pearmain is a an old-fashioned late dessert apple, one of the most popular varieties in Victorian England. It has an attractive 'pearmain' shape. This is a fairly dry apple - which is perhaps not regarded as a desirable attribute today. In spite of this it is actually a very enjoyable apple, with a rich aromatic flavour which in apple terms is usually described as Although it had 'shelf appeal' for the Victorian housewife, its autumnal colouring is probably too subdued to compete with the bright young things of the modern supermarket shelves. Perhaps this is part of its appeal; it recalls a bygone era where subtlety of flavour was appreciated - a lovely apple to savour in front of an open fire on a cold winter's day. Tree hardy. Does will in all soils, even clay. AERLIE RED FLESH (Hidden Rose, Mountain Rose) California 1930’s 19 20 20 Cook Oct 20 15 An amazing red fleshed apple, discovered in Aerlie, Oregon, which may be the best of all red fleshed varieties and indeed would be an outstandingly delicious apple no matter what color the flesh is. A choice seedling, Aerlie Red Flesh has a beautiful yellow skin with pale whitish dots, but it is inside that it excels. Deep rose red flesh, juicy, crisp, hard, sugary and richly flavored, ripening late (October) and keeping throughout the winter. The late Conrad Gemmer, an astute observer of apples with 500 varieties in his collection, rated Hidden Rose an outstanding variety of top quality. -

APPLE (Fruit Varieties)
E TG/14/9 ORIGINAL: English DATE: 2005-04-06 INTERNATIONAL UNION FOR THE PROTECTION OF NEW VARIETIES OF PLANTS GENEVA * APPLE (Fruit Varieties) UPOV Code: MALUS_DOM (Malus domestica Borkh.) GUIDELINES FOR THE CONDUCT OF TESTS FOR DISTINCTNESS, UNIFORMITY AND STABILITY Alternative Names:* Botanical name English French German Spanish Malus domestica Apple Pommier Apfel Manzano Borkh. The purpose of these guidelines (“Test Guidelines”) is to elaborate the principles contained in the General Introduction (document TG/1/3), and its associated TGP documents, into detailed practical guidance for the harmonized examination of distinctness, uniformity and stability (DUS) and, in particular, to identify appropriate characteristics for the examination of DUS and production of harmonized variety descriptions. ASSOCIATED DOCUMENTS These Test Guidelines should be read in conjunction with the General Introduction and its associated TGP documents. Other associated UPOV documents: TG/163/3 Apple Rootstocks TG/192/1 Ornamental Apple * These names were correct at the time of the introduction of these Test Guidelines but may be revised or updated. [Readers are advised to consult the UPOV Code, which can be found on the UPOV Website (www.upov.int), for the latest information.] i:\orgupov\shared\tg\applefru\tg 14 9 e.doc TG/14/9 Apple, 2005-04-06 - 2 - TABLE OF CONTENTS PAGE 1. SUBJECT OF THESE TEST GUIDELINES..................................................................................................3 2. MATERIAL REQUIRED ...............................................................................................................................3 -

2019 Newsletter
Front page: Allen’s greeting, something new 2019 NEWSLETTER A Message From Our President & Owner, EVENT CALENDAR Cooler mornings and valley fog below the orchard remind us all that it’s about apple time! Nature has blessed us with August 19th a beautiful crop of apples with exceptionally good fruit size. Opening Day Compared to recent years, some varieties may be picked a little later this year so be sure to give us a call or check our website to September 27th - 29th make sure your favorite apple is available. I enjoy every apple Gays Mills Apple Festival variety we grow, but Evercrisp has me as excited as Honeycrisp. October 5th - 6th Harvested in late October and stored in a refrigerator, Evercrisp Sunrise Samples Weekend is a fantastic eating experience in the winter months. Our family has been growing apples since 1934 and we have never tasted October 12th - 13th another winter apple like Evercrisp! Family Fun Weekend I hope you all enjoyed our newly expanded sales area and October 19th - 20th bathrooms added in 2018. This year we have made additional Harvest Celebration exciting improvements with a new gift area, live apple packing & Helicopter Rides TV, and a working model train for young and old to enjoy. Our famous cider donuts will be back- made fresh every day. Please (weather permitting ) enjoy our free apple and cider samples along with many of the October 21st - December 16th other products we sell. Gift Box Shipping Begins Don’t forget our online store. We feature many of the October 26th - 27th items available here and have made it far easier to order gift pack Trick or Treat Weekend apples this year from home. -

Variety Description Origin Approximate Ripening Uses
Approximate Variety Description Origin Ripening Uses Yellow Transparent Tart, crisp Imported from Russia by USDA in 1870s Early July All-purpose Lodi Tart, somewhat firm New York, Early 1900s. Montgomery x Transparent. Early July Baking, sauce Pristine Sweet-tart PRI (Purdue Rutgers Illinois) release, 1994. Mid-late July All-purpose Dandee Red Sweet-tart, semi-tender New Ohio variety. An improved PaulaRed type. Early August Eating, cooking Redfree Mildly tart and crunchy PRI release, 1981. Early-mid August Eating Sansa Sweet, crunchy, juicy Japan, 1988. Akane x Gala. Mid August Eating Ginger Gold G. Delicious type, tangier G Delicious seedling found in Virginia, late 1960s. Mid August All-purpose Zestar! Sweet-tart, crunchy, juicy U Minn, 1999. State Fair x MN 1691. Mid August Eating, cooking St Edmund's Pippin Juicy, crisp, rich flavor From Bury St Edmunds, 1870. Mid August Eating, cider Chenango Strawberry Mildly tart, berry flavors 1850s, Chenango County, NY Mid August Eating, cooking Summer Rambo Juicy, tart, aromatic 16th century, Rambure, France. Mid-late August Eating, sauce Honeycrisp Sweet, very crunchy, juicy U Minn, 1991. Unknown parentage. Late Aug.-early Sept. Eating Burgundy Tart, crisp 1974, from NY state Late Aug.-early Sept. All-purpose Blondee Sweet, crunchy, juicy New Ohio apple. Related to Gala. Late Aug.-early Sept. Eating Gala Sweet, crisp New Zealand, 1934. Golden Delicious x Cox Orange. Late Aug.-early Sept. Eating Swiss Gourmet Sweet-tart, juicy Switzerland. Golden x Idared. Late Aug.-early Sept. All-purpose Golden Supreme Sweet, Golden Delcious type Idaho, 1960. Golden Delicious seedling Early September Eating, cooking Pink Pearl Sweet-tart, bright pink flesh California, 1944, developed from Surprise Early September All-purpose Autumn Crisp Juicy, slow to brown Golden Delicious x Monroe. -

Cosmic Crisp's Growth Is out of This World
- Advertisement - Cosmic Crisp's growth is out of this world 1 / 2 April 26, 2021 After a record-breaking sophomore year that hasn't finished, the Cosmic Crisp apple’s growth trajectory indicates it will launch to the top of the apple chart in no time. Nielsen data show monumental growth, reaching No. 11 in sales dollars in March, and No. 14 over the 52 weeks ending March 27. If retailers do not have a Cosmic Crisp apple program in place now, they are missing out on very important sales and customer expectations. “Cosmic Crisp volume is planned to more than double this fall, and double again for the 2022 harvest. This volume will rival current core variety volume, displacing mature varieties that are being replaced with higher color strains and higher flavor varieties,” said Catherine Gipe-Stewart, communications manager. “Cosmic Crisp is rising so quickly, it has potential to become a top-five variety at this time next year.” Nielsen data show a 595 percent increase in dollars and a 720 percent in volume over the four weeks ending March 27. In March alone, Cosmic Crisp earned the 11th spot with $4.3 million in sales and 1.8 million pounds, at an average price of $2.52 per pound, right in line with Honeycrisp at $2.55 per pound. Looking at a yearly perspective (last 52 weeks), Cosmic Crisp has earned $20 million in sales, with 7.7 million pounds. “Historically, apples like Cosmic Crisp take root in the Pacific region, and spread eastward like wildfire,” said Gipe-Stewart. -

Università Degli Studi Di Palermo
UNIVERSITÀ DEGLI STUDI DI PALERMO Dottorato Scienze Agrarie Forestali e Ambientali Dipartimento Scienze Agrarie, Alimentari e Forestali Settore Scientifico Disciplinare AGR/03 INVESTIGATION OF BIOTIC STRESS RESPONSES IN FRUIT TREE CROPS USING META-ANALYTICAL TECHNIQUES IL DOTTORE IL COORDINATORE BIPIN BALAN CH.MO PROF. VINCENZO BAGARELLO IL TUTOR CO TUTOR CH.MO PROF. TIZIANO CARUSO CH.MO PROF. FEDERICO MARTINELLI CICLO XXXII ANNO CONSEGUIMENTO TITOLO 2020 Table of Contents State of art …...………………………………………………………….......…………6 Experiment 1……..…………………….…...………………………….......……..…10 Transcriptomic responses to biotic stresses in Malus x domestica: a meta-analysis study. ………………………….……………………....................................……..…10 1. Introduction ………………………………………………...................….……..…10 2. Materials and methods ……………………………………..………….....…..……21 2.1 Search strategy of published study identification and selection for meta- analysis …………………………………………………………........….……21 2.2 Extraction and annotation of differentially expressed genes ………..…....23 2.3 Gene enrichment analysis ……………………………………….......……23 2.4 Functional analysis…………………………………..……….......….……23 2.5 Protein-protein interaction network……………………..……….……..…25 3. Results……...…………………………………….....................................……...…27 3.1 Meta-analysis of transcriptome data ……………………….........……..…27 3.2 Gene ontology analysis …………………………………………….…..…28 3.3 Gene set enrichment analysis …………………………………....……..…32 3.4 Metabolism overview ………………………………………………….…34 3.5 Hormone-related pathways ……………………………………...……..…35 -

Survey of Apple Clones in the United States
Historic, archived document Do not assume content reflects current scientific knowledge, policies, or practices. 5 ARS 34-37-1 May 1963 A Survey of Apple Clones in the United States u. S. DFPT. OF AGRffini r U>2 4 L964 Agricultural Research Service U.S. DEPARTMENT OF AGRICULTURE PREFACE This publication reports on surveys of the deciduous fruit and nut clones being maintained at the Federal and State experiment stations in the United States. It will b- published in three c parts: I. Apples, II. Stone Fruit. , UI, Pears, Nuts, and Other Fruits. This survey was conducted at the request of the National Coor- dinating Committee on New Crops. Its purpose is to obtain an indication of the volume of material that would be involved in establishing clonal germ plasm repositories for the use of fruit breeders throughout the country. ACKNOWLEDGMENT Gratitude is expressed for the assistance of H. F. Winters of the New Crops Research Branch, Crops Research Division, Agricultural Research Service, under whose direction the questionnaire was designed and initial distribution made. The author also acknowledges the work of D. D. Dolan, W. R. Langford, W. H. Skrdla, and L. A. Mullen, coordinators of the New Crops Regional Cooperative Program, through whom the data used in this survey were obtained from the State experiment stations. Finally, it is recognized that much extracurricular work was expended by the various experiment stations in completing the questionnaires. : CONTENTS Introduction 1 Germany 298 Key to reporting stations. „ . 4 Soviet Union . 302 Abbreviations used in descriptions .... 6 Sweden . 303 Sports United States selections 304 Baldwin. -

Assessment of Allelic Diversity in Intron-Containing Mal D 1 Genes and Their
BMC Plant Biology BioMed Central Research article Open Access Assessment of allelic diversity in intron-containing Mal d 1 genes and their association to apple allergenicity Zhongshan Gao1,2,3, Eric W van de Weg*2,3, Catarina I Matos2, Paul Arens3, Suzanne THP Bolhaar4, Andre C Knulst4, Yinghui Li3,5, Karin Hoffmann- Sommergruber6 and Luud JWJ Gilissen2,3 Address: 1Department of Horticulture/Allergy Research Center, Zhejiang University, Hangzhou 310029, PR China, 2Allergy Consortium Wageningen, Wageningen University and Research Centre, P.O. Box 16, 6700AA, Wageningen, the Netherlands, 3Plant Research International, Wageningen University and Research Centre, P.O. Box 16, 6700AA, Wageningen, the Netherlands, 4Department of Dermatology/Allergology, University Medical Center Utrecht, P.O. Box 85500, 3508GA Utrecht, the Netherlands, 5The National Key Facility for Crop Gene Resources and Genetic Improvement (NFCRI)/Key Lab of Germplasm & Biotechnology (MOA), Institute of Crop Science, China Academy of Agricultural Science, Beijing, 100081, PR China and 6Department of Pathophysiology, Medical University of Vienna, AKH-EBO-3Q, Währinger Gürtel 18-20, A-1090 Vienna, Austria Email: Zhongshan Gao - [email protected]; Eric W van de Weg* - [email protected]; Catarina I Matos - [email protected]; Paul Arens - [email protected]; Suzanne THP Bolhaar - [email protected]; Andre C Knulst - [email protected]; Yinghui Li - [email protected]; Karin Hoffmann-Sommergruber - Karin.Hoffmann- [email protected]; Luud JWJ Gilissen - [email protected] * Corresponding author Published: 13 November 2008 Received: 28 November 2007 Accepted: 13 November 2008 BMC Plant Biology 2008, 8:116 doi:10.1186/1471-2229-8-116 This article is available from: http://www.biomedcentral.com/1471-2229/8/116 © 2008 Gao et al; licensee BioMed Central Ltd. -

Handling of Apple Transport Techniques and Efficiency Vibration, Damage and Bruising Texture, Firmness and Quality
Centre of Excellence AGROPHYSICS for Applied Physics in Sustainable Agriculture Handling of Apple transport techniques and efficiency vibration, damage and bruising texture, firmness and quality Bohdan Dobrzañski, jr. Jacek Rabcewicz Rafa³ Rybczyñski B. Dobrzañski Institute of Agrophysics Polish Academy of Sciences Centre of Excellence AGROPHYSICS for Applied Physics in Sustainable Agriculture Handling of Apple transport techniques and efficiency vibration, damage and bruising texture, firmness and quality Bohdan Dobrzañski, jr. Jacek Rabcewicz Rafa³ Rybczyñski B. Dobrzañski Institute of Agrophysics Polish Academy of Sciences PUBLISHED BY: B. DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES ACTIVITIES OF WP9 IN THE CENTRE OF EXCELLENCE AGROPHYSICS CONTRACT NO: QLAM-2001-00428 CENTRE OF EXCELLENCE FOR APPLIED PHYSICS IN SUSTAINABLE AGRICULTURE WITH THE th ACRONYM AGROPHYSICS IS FOUNDED UNDER 5 EU FRAMEWORK FOR RESEARCH, TECHNOLOGICAL DEVELOPMENT AND DEMONSTRATION ACTIVITIES GENERAL SUPERVISOR OF THE CENTRE: PROF. DR. RYSZARD T. WALCZAK, MEMBER OF POLISH ACADEMY OF SCIENCES PROJECT COORDINATOR: DR. ENG. ANDRZEJ STĘPNIEWSKI WP9: PHYSICAL METHODS OF EVALUATION OF FRUIT AND VEGETABLE QUALITY LEADER OF WP9: PROF. DR. ENG. BOHDAN DOBRZAŃSKI, JR. REVIEWED BY PROF. DR. ENG. JÓZEF KOWALCZUK TRANSLATED (EXCEPT CHAPTERS: 1, 2, 6-9) BY M.SC. TOMASZ BYLICA THE RESULTS OF STUDY PRESENTED IN THE MONOGRAPH ARE SUPPORTED BY: THE STATE COMMITTEE FOR SCIENTIFIC RESEARCH UNDER GRANT NO. 5 P06F 012 19 AND ORDERED PROJECT NO. PBZ-51-02 RESEARCH INSTITUTE OF POMOLOGY AND FLORICULTURE B. DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES ©Copyright by BOHDAN DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES LUBLIN 2006 ISBN 83-89969-55-6 ST 1 EDITION - ISBN 83-89969-55-6 (IN ENGLISH) 180 COPIES, PRINTED SHEETS (16.8) PRINTED ON ACID-FREE PAPER IN POLAND BY: ALF-GRAF, UL. -
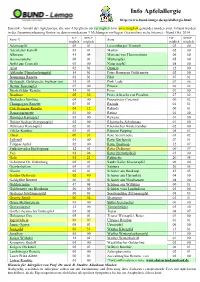
02 Apfelallergie Sortenliste Allgemein
Info Apfelallergie http://www.bund-lemgo.de/apfelallergie.html Statistik - Anzahl der Apfelsorten, die von Allergikern als verträglich bzw. unverträglich gemeldet worden sind. Erfasst werden in der Zusammenfassung Sorten zu denen mindestens 3 Meldungen vorliegen. Gesamtliste siehe Internet - Stand Okt. 2014 ver- unver- ver- unver- Sorte *) Sorte träglich träglich träglich träglich Adamsapfel 05 01 Luxemburger Triumph 03 00 Adersleber Kalvill 03 01 Martini 02 00 Alkmene 41 04 Minister von Hammerstein 06 00 Ananasrenette 06 01 Mutterapfel 05 00 Apfel aus Croncels 03 00 Notarisapfel 08 00 Arlet 02 01 Ontario 12 00 Altländer Pfannkuchenapfel 14 01 Peter Heusgens Goldrenette 02 00 Baumanns Renette 01 01 Pilot 07 01 Berlepsch, Goldrenette Freiherr von 35 03 Pink Lady 07 06 Berner Rosenapfel 07 00 Pinova 06 02 Biesterfelder Renette 14 01 Piros 01 00 Braeburn 05 32 Prinz Albrecht von Preußen 27 02 Brettacher Sämling 04 00 Purpurroter Cousinot 00 02 Champagner Renette 07 03 Reanda 01 01 Cox Orangen-Renette 04 12 Relinda 00 01 Damasonrenette 03 00 Retina 01 00 Danziger Kantapfel 05 00 Rewena 01 00 Doktor Seeligs Orangenapfel 02 00 Rheinische Schafsnase 01 00 Dülmener Rosenapfel 02 01 Rheinischer Winterrambur 02 00 Eifeler Rambur 03 01 Ripston Pepping 05 01 Elstar 09 31 Rote Sternrenette 03 02 Fallstaff 01 00 Roter Berlepsch 07 01 Filippas Apfel 02 00 Roter Boskoop 32 07 Finkenwerder Herbstprinz 12 01 Roter Delicious 00 07 Fuji 01 06 Roter Herbstkalvill 03 00 Gala 01 21 Rubinette 20 04 Geheimrat Dr. Oldenburg 02 03 Sankt Galler Klosterapfel 06 00 -

Poems of John Donne
1/ THE LIBRARY OF THE UNIVERSITY OF NORTH CAROLINA ENDOWED BY THE DIALECTIC AND PHILANTHROPIC SOCIETIES PR22U5 .A5 C5 1901, v. 2 UNIVERSITY OF N.C. AT CHAPEL HILL 10001490423 This book is due at the LOUIS R. WILSON LIBRARY on the last date stamped under "Date Due." If not on hold it may be renewed by bringing it to the library. DATE DS. T DATE DUE RET DUE KL1 '1 fit o n tHGf ..1. _ r\ *C\ n \j n f 1 All -"\ a« »/*V JAN23 9! 1 1 7QQ2 ArK V) r POEMS OF JOHN DONNE. Digitized by the Internet Archive in 2014 https://archive.org/details/poemsofjohndonne02donn_0 POEMS .tf** C 6~ OF )*0 I JOHN DONNE^ E. K. CHAMBERS. WITH AN INTRODUCTION BY GEORGE S AINTSBURY. NEW EDITION. VOL. II. LONDON: NEW YORK: A. H. BTJLLEN, CHARLES SCRIBNER'S SONS, 18 Cecil Court, W.C. i53~7 Fifth Avenue. xgoj^ 1901. Richard Clay &,Sons, Limited, London & Bungay. CONTENTS OF VOL. II. PAGE Table of Contents ... vii Letters to Several Personages— To Mr. Christopher Brooke : The Storm I „ „ „ The Calm 4 To Sir Henry Wotton ... 7 To Sir Henry Goodyere 10 To Mr. Rowland Woodward .. ... 12 To Sir Henry Wotton 14 To the Countess of Bedford 15 To the Countess of Bedford 17 To Sir Edward Herbert. 20 To the Countess of Bedford 22 To the Countess of Bedford, on New Year's Day 26 To the Countess of Huntingdon ... ... 29 To M[r] Ifzaak] W[alton] ... 32 To M[r] T. W 33 To M[r] T.