Spatial Predictive Distribution Modelling of Madeira's Endemic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

SPIRULA - Speciale Uitgave, Supplement Nr
SPIRULA - Speciale uitgave, supplement nr. 2 13 Eponiemen betreffende personen verbonden aan de NMV Gijs+C. Kronenberg Inleiding In het kader van het 50-jarig jubileum van de NMV werd een lijst gepubliceerd (KUIPER, 1984: 1589-1590) met In eponiemen, dat wil zeggen, weekdiersoortenvernoemd naar Nederlandse malacologen. latere jaren verschenen hierop een aantal aanvullingen (KUIPER, 1986; KUIPER, 1988; KUIPER, 1989; KUIPER, 1991; KUIPER, 1992; KUIPER, 1993; KUIPER 1995). Na 1995 zijn er verder geen overzichten meer samengesteld. Helaas is aan het initiatiefvan met KUIPER (opera cit.) geen vervolg gegeven, zodat we een achterstand zitten, maar die nu wordt weggewerkt middels deze vernieuwde lijst. Criteria perd door [KRONENBERG] (2001). Om in de lijst opgenomen te worden dientte worden voldaan Daarnaast hebben deze criteriaook als gevolg dat de namen aan drie criteria: Trochus wilsi PICKERY, 1989 (KUIPER 1992); Ischnochiton Men moet lid zijn (geweest) van de N.M.V. ofeen plaatselij- vanbellei KAAS; en Notoplax richardi KAAS [ook vernoemd ke schelpenwerkgroep enigszins geliëerd aan de NMV, of naar de heer R.A. VAN BELLE] (KUIPER, 1993) uitde lijst zijn daar in zeer nauwe (privé) relatie mee staan (huwelijk, geschrapt daar deze personen niet de Nederlandse nationali- dank samenwonen, kinderen) als voor de vele jaren trouwe teit (gehad) hebben, en taxa vernoemd naar Prof. Dr. J.K.L. Of in niet daar deze ondersteuning en dergelijke. men moet dusdanige pro- MARTIN opgenomen zijn niet de Nederlandse fessioneel malacologische relatie hebben gestaan dat de nationaliteitheeft gehad. auteur het taxon dat tot wilde in het Taxa beschreven door b.v. uit van uiting brengen ver- E.J. -

Diplom-Biologe KLAUS GROH Malakozoologe Und Naturschützer – 65 Jahre
53 Mitt. dtsch. malakozool. Ges. 94 53 – 70 Frankfurt a. M., November 2015 Diplom-Biologe KLAUS GROH Malakozoologe und Naturschützer – 65 Jahre CARSTEN RENKER & JÜRGEN H. JUNGBLUTH th Abstract: The 65 birthday of KLAUS GROH is a good occasion to give a retrospect of his life and hitherto existing achievement. Beside his vita we summarize his malacological work, give an overview about the projects for the protection of species, have a look on his tremendous impetus for the worldwide distribution of malacological knowledge by the establishment of the CHRISTA HEMMEN-Verlag, later ConchBooks, as publishing house, book trader and antiquarian. Last but not least we give a summary of his scientific achievements culminating in 206 publications and containing descriptions of up to now 42 specific taxa. Keywords: KLAUS GROH, biography, bibliography, malacology, freshwater mussels, Hesse, Rhineland- Palatinate, Luxembourg Zusammenfassung: Der 65. Geburtstag von KLAUS GROH wird zum Anlass genommen einen Rückblick auf sein bisheriges Leben und Wirken zu geben. Neben der Vita werden vor allem seine malakologische Arbeit und sein ehrenamtliches Engagement in zahlreichen malakologischen Verbänden und Naturschutzvereinen betrachtet. KLAUS GROH nahm außerdem einen enormen Einfluss auf die weltweite Verbreitung malako- logischen Wissens durch die Gründung des CHRISTA HEMMEN-Verlags, später ConchBooks, als Verlagshaus, Buchhandlung und Antiquariat. Schließlich gilt es seine wissenschaftlichen Verdienste zu würdigen, die in 206 Publikationen und Neubeschreibungen 42 spezifischer Taxa kulminieren. Vita Schulzeit Am 22. Mai 1949 wurde KLAUS GROH in Darmstadt als Sohn des Bauschlossers HELMUT GROH und seiner Ehefrau ANNELIESE, geb. FEDERLEIN geboren. Er besuchte die Volksschulen in Langen/Hessen und Kirchheim unter Teck/Baden-Württemberg (1955-1959), es folgte der Besuch der Realschule in Langen/Hessen (1959-1965), dort schloss er auch seine Schulzeit mit der „Mittleren Reife“ ab. -
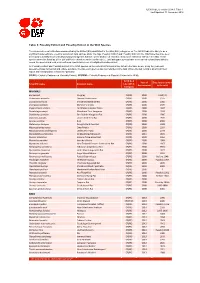
Table 9: Possibly Extinct and Possibly Extinct in the Wild Species
IUCN Red List version 2014.3: Table 9 Last Updated: 13 November 2014 Table 9: Possibly Extinct and Possibly Extinct in the Wild Species The number of recent extinctions documented by the Extinct (EX) and Extinct in the Wild (EW) categories on The IUCN Red List is likely to be a significant underestimate, even for well-known taxa such as birds. The tags 'Possibly Extinct' and 'Possibly Extinct in the Wild' have therefore been developed to identify those Critically Endangered species that are, on the balance of evidence, likely to be extinct (or extinct in the wild). These species cannot be listed as EX or EW until their extinction can be confirmed (i.e., until adequate surveys have been carried out and have failed to record the species and local or unconfirmed reports have been investigated and discounted). All 'Possibly Extinct' and 'Possibly Extinct in the Wild' species on the current IUCN Red List are listed in the table below, along the year each assessment was carried out and, where available, the date each species was last recorded in the wild. Where the last record is an unconfirmed report, last recorded date is noted as "possibly". CR(PE) - Critically Endangered (Possibly Extinct), CR(PEW) - Critically Endangered (Possibly Extinct in the Wild), IUCN Red Year of Date last recorded Scientific name Common name List (2014) Assessment in the wild Category MAMMALS Bos sauveli Kouprey CR(PE) 2008 1969/70 Crateromys australis Dinagat Crateromys CR(PE) 2008 1975 Crocidura trichura Christmas Island Shrew CR(PE) 2008 1985 Crocidura wimmeri -

Fauna of New Zealand Ko Te Aitanga Pepeke O Aotearoa
aua o ew eaa Ko te Aiaga eeke o Aoeaoa IEEAE SYSEMAICS AISOY GOU EESEAIES O ACAE ESEAC ema acae eseac ico Agicuue & Sciece Cee P O o 9 ico ew eaa K Cosy a M-C aiièe acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa EESEAIE O UIESIIES M Emeso eame o Eomoogy & Aima Ecoogy PO o ico Uiesiy ew eaa EESEAIE O MUSEUMS M ama aua Eiome eame Museum o ew eaa e aa ogaewa O o 7 Weigo ew eaa EESEAIE O OESEAS ISIUIOS awece CSIO iisio o Eomoogy GO o 17 Caea Ciy AC 1 Ausaia SEIES EIO AUA O EW EAA M C ua (ecease ue 199 acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa Fauna of New Zealand Ko te Aitanga Pepeke o Aotearoa Number / Nama 38 Naturalised terrestrial Stylommatophora (Mousca Gasooa Gay M ake acae eseac iae ag 317 amio ew eaa 4 Maaaki Whenua Ρ Ε S S ico Caeuy ew eaa 1999 Coyig © acae eseac ew eaa 1999 o a o is wok coee y coyig may e eouce o coie i ay om o y ay meas (gaic eecoic o mecaica icuig oocoyig ecoig aig iomaio eiea sysems o oewise wiou e wie emissio o e uise Caaoguig i uicaio AKE G Μ (Gay Micae 195— auase eesia Syommaooa (Mousca Gasooa / G Μ ake — ico Caeuy Maaaki Weua ess 1999 (aua o ew eaa ISS 111-533 ; o 3 IS -7-93-5 I ie 11 Seies UC 593(931 eae o uIicaio y e seies eio (a comee y eo Cosy usig comue-ase e ocessig ayou scaig a iig a acae eseac M Ae eseac Cee iae ag 917 Aucka ew eaa Māoi summay e y aco uaau Cosuas Weigo uise y Maaaki Weua ess acae eseac O o ico Caeuy Wesie //wwwmwessco/ ie y G i Weigo o coe eoceas eicuaum (ue a eigo oaa (owe (IIusao G M ake oucio o e coou Iaes was ue y e ew eaIa oey oa ue oeies eseac -

Amaia Caro Aramendia
The genus Pyrenaearia (Gastropoda, Helicoidea): Molecular and Morphological Systematics, Biogeography and Population Dynamics Pyrenaearia generoa (Gastropoda, Helicoidea): Sistematika Molekularra eta Morfologikoa, Biogeografia eta Populazio Dinamika PhD thesis Vitoria-Gasteiz, 2019 Amaia Caro Aramendia The genus Pyrenaearia (Gastropoda, Helicoidea): Molecular and Morphological Systematics, Biogeography and Population Dynamics Pyrenaearia generoa (Gastropoda, Helicoidea): Sistematika Molekularra eta Morfologikoa, Biogeografia eta Populazio Dinamika A thesis submitted by Amaia Caro Aramendia for the degree of Doctor of Philosophy, under the supervision of Dr. Benjamín Juan Gómez-Moliner and Dr. María José Madeira University of the Basque Country, Vitoria-Gasteiz, 2019 Zoologia eta Animalia Biologia Zelulen Saila Dpto. Zoología y Biología Celular Animal (cc)2019 AMAIA CARO ARAMENDIA (cc by-nc-nd 4.0) Astiro igo, barraskilotxo Fuji mendia da hau! Kobayashi Issa-ren haikua To the little things that run the world Esker onak Acknowledgements Tesi bat ez da pertsona bakar batena, bidean zehar laguntzen duten pertsona guztiei esker sortutako lana da eta, beraz, lehen orriek haien laguntza eskertzeko izan behar dute: En primer lugar me gustaría agradecer a mis directores, Benjamín Gómez-Moliner y María José Madeira. A Benjamín, por darme la oportunidad de entrar en el grupo de investigación y confiar en que podría realizar esta tesis. Gracias por compartir tus extensos conocimientos y por descubrirme el mundo de la malacología, que sin duda no habría encontrado por mi cuenta y ha resultado de lo más interesante. A Marijo, porque desde el principio y hasta el final has estado siempre ahí para guiarme, animarme y para ayudarme en todo lo que hiciese falta pero, sobre todo, por mostrarme que es posible compaginar este trabajo con una vida fuera de él. -

(Approx) Mixed Micro Shells (22G Bags) Philippines € 10,00 £8,64 $11,69 Each 22G Bag Provides Hours of Fun; Some Interesting Foraminifera Also Included
Special Price £ US$ Family Genus, species Country Quality Size Remarks w/o Photo Date added Category characteristic (€) (approx) (approx) Mixed micro shells (22g bags) Philippines € 10,00 £8,64 $11,69 Each 22g bag provides hours of fun; some interesting Foraminifera also included. 17/06/21 Mixed micro shells Ischnochitonidae Callistochiton pulchrior Panama F+++ 89mm € 1,80 £1,55 $2,10 21/12/16 Polyplacophora Ischnochitonidae Chaetopleura lurida Panama F+++ 2022mm € 3,00 £2,59 $3,51 Hairy girdles, beautifully preserved. Web 24/12/16 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 30mm+ € 4,00 £3,45 $4,68 30/04/21 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 27.9mm € 2,80 £2,42 $3,27 30/04/21 Polyplacophora Ischnochitonidae Stenoplax limaciformis Panama F+++ 16mm+ € 6,50 £5,61 $7,60 Uncommon. 24/12/16 Polyplacophora Chitonidae Acanthopleura gemmata Philippines F+++ 25mm+ € 2,50 £2,16 $2,92 Hairy margins, beautifully preserved. 04/08/17 Polyplacophora Chitonidae Acanthopleura gemmata Australia F+++ 25mm+ € 2,60 £2,25 $3,04 02/06/18 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 41mm+ € 4,00 £3,45 $4,68 West Indian 'fuzzy' chiton. Web 24/12/16 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 32mm+ € 3,00 £2,59 $3,51 West Indian 'fuzzy' chiton. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 44mm+ € 5,00 £4,32 $5,85 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F++ 35mm € 2,50 £2,16 $2,92 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 29mm+ € 3,00 £2,59 $3,51 Caribbean. -

Malacologica
FOLIA Folia Malacol. 24(3): 111–177 MALACOLOGICA ISSN 1506-7629 The Association of Polish Malacologists Faculty of Biology, Adam Mickiewicz University Bogucki Wydawnictwo Naukowe Poznań, September 2016 http://dx.doi.org/10.12657/folmal.024.008 PATTERNS OF SPATIO-TEMPORAL VARIATION IN LAND SNAILS: A MULTI-SCALE APPROACH SERGEY S. KRAMARENKO Mykolaiv National Agrarian University, Paryzka Komuna St. 9, Mykolaiv, 54020, Ukraine (e-mail: [email protected]) ABSTRACT: Mechanisms which govern patterns of intra-specific vatiation in land snails were traced within areas of different size, using Brephulopsis cylindrica (Menke), Chondrula tridens (O. F. Müller), Xeropicta derbentina (Krynicki), X. krynickii (Krynicki), Cepaea vindobonensis (Férussac) and Helix albescens Rossmässler as examples. Morphometric shell variation, colour and banding pattern polymorphism as well as genetic polymorphism (allozymes and RAPD markers) were studied. The results and literature data were analysed in an attempt to link patterns to processes, with the following conclusions. Formation of patterns of intra- specific variation (initial processes of microevolution) takes different course at three different spatial scales. At micro-geographical scale the dominant role is played by eco-demographic characteristics of the species in the context of fluctuating environmental factors. At meso-geographical scale a special part is played by stochastic population-genetic processes. At macro-geographical scale more or less distinct clinal patterns are associated with basic macroclimatic -
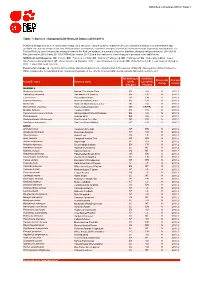
Table 7: Species Changing IUCN Red List Status (2010-2011)
IUCN Red List version 2011.2: Table 7 Table 7: Species changing IUCN Red List Status (2010-2011) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2010 (IUCN Red List version 2010.4) and 2011 (IUCN Red List version 2011.2) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.) IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2010) List (2011) change version Category Category MAMMALS Bradypus torquatus Maned Three-toed Sloth EN VU N 2011.1 Callicebus oenanthe San Martin Titi Monkey EN CR N 2011.1 Equus ferus Przewalski's Horse CR EN G 2011.2 -

Organismu Latviskie Nosaukumi (2)
Biosistēmu Terminoloģijas Centra Biļetens 1(1) (2017): 21–51 ISSN 2501-0336 (online) http://www.rpd-science.org/BTCB/V001/BTCB_1_4.pdf © “RPD Science” Citēšanai: BTCB, 2017. Organismu latviskie nosaukumi (2). Biosistēmu Terminoloģijas Centra Biļetens 1(1): 21–51 Organismu latviskie nosaukumi (2) Latvian names of organisms (2) Zinātniskais nosaukums Atbilstība Pēdējā Scientific name Equivalence pārbaude Last verification A Abies gmelinii Rupr. (1845) = Larix gmelinii (Rupr.) Rupr. var. gmelinii 17.12.2016. Abies ledebourii Rupr. (1845) = Larix gmelinii (Rupr.) Rupr. var. gmelinii 17.12.2016. Abies menziesii Mirb. (1825) = Pseudotsuga menziesii (Mirb.) Franco var. menziesii 17.12.2016. Acaciaceae = Fabaceae 27.12.2016. Acalitus brevitarsus (Fockeu, 1890) melnalkšņa maurērce 16.12.2016. Acalitus calycophthirus (Nalepa, 1891) bērzu pumpurērce 16.12.2016. Acalitus essigi (Hassan, 1928) aveņu pangērce 16.12.2016. Acalitus longisetosus (Nalepa, 1892) bērzu sārtā maurērce 16.12.2016. Acalitus phloeocoptes (Nalepa, 1890) plūmju stumbra pangērce 16.12.2016. Acalitus phyllereus (Nalepa, 1919) baltalkšņa maurērce 16.12.2016. Acalitus plicans (Nalepa, 1917) dižskābaržu maurērce 16.12.2016. Acalitus rudis (Canestrini, 1890) bērzu baltā maurērce 16.12.2016. Acalitus stenaspis (Nalepa, 1891) dižskābaržu lapmalērce 16.12.2016. Acalitus vaccinii (Keifer, 1939) melleņu pumpurērce 16.12.2016. Acanthinula aculeata (O. F. Müller, 1774) mazais dzeloņgliemezis 16.12.2016. Acanthinula spinifera Mousson, 1872 Spānijas dzeloņgliemezis 16.12.2016. Acanthocardia echinata (Linnaeus, 1758) dzelkņainā sirsniņgliemene 16.12.2016. Acanthochitona crinita (Pennant, 1777) zaļais bruņgliemis 16.12.2016. Aceria brevipunctatus (Nalepa, 1889) = Aceria campestricola (Frauenfeld, 1865) 16.12.2016. Aceria brevirostris (Nalepa, 1892) ziepenīšu pangērce 16.12.2016. Aceria brevitarsus (Fockeu, 1890) = Acalitus brevitarsus (Fockeu, 1890) 16.12.2016. -

Fauna of New Zealand Website Copy 2010, Fnz.Landcareresearch.Co.Nz
aua o ew eaa Ko te Aiaga eeke o Aoeaoa IEEAE SYSEMAICS AISOY GOU EESEAIES O ACAE ESEAC ema acae eseac ico Agicuue & Sciece Cee P O o 9 ico ew eaa K Cosy a M-C aiièe acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa EESEAIE O UIESIIES M Emeso eame o Eomoogy & Aima Ecoogy PO o ico Uiesiy ew eaa EESEAIE O MUSEUMS M ama aua Eiome eame Museum o ew eaa e aa ogaewa O o 7 Weigo ew eaa EESEAIE O OESEAS ISIUIOS awece CSIO iisio o Eomoogy GO o 17 Caea Ciy AC 1 Ausaia SEIES EIO AUA O EW EAA M C ua (ecease ue 199 acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa Fauna of New Zealand Ko te Aitanga Pepeke o Aotearoa Number / Nama 38 Naturalised terrestrial Stylommatophora (Mousca Gasooa Gay M ake acae eseac iae ag 317 amio ew eaa 4 Maaaki Whenua Ρ Ε S S ico Caeuy ew eaa 1999 Coyig © acae eseac ew eaa 1999 o a o is wok coee y coyig may e eouce o coie i ay om o y ay meas (gaic eecoic o mecaica icuig oocoyig ecoig aig iomaio eiea sysems o oewise wiou e wie emissio o e uise Caaoguig i uicaio AKE G Μ (Gay Micae 195— auase eesia Syommaooa (Mousca Gasooa / G Μ ake — ico Caeuy Maaaki Weua ess 1999 (aua o ew eaa ISS 111-533 ; o 3 IS -7-93-5 I ie 11 Seies UC 593(931 eae o uIicaio y e seies eio (a comee y eo Cosy usig comue-ase e ocessig ayou scaig a iig a acae eseac M Ae eseac Cee iae ag 917 Aucka ew eaa Māoi summay e y aco uaau Cosuas Weigo uise y Maaaki Weua ess acae eseac O o ico Caeuy Wesie //wwwmwessco/ ie y G i Weigo o coe eoceas eicuaum (ue a eigo oaa (owe (IIusao G M ake oucio o e coou Iaes was ue y e ew eaIa oey oa ue oeies eseac -

Leiostyla L Ove, 1852 Mollusca, Pulmonata Новые Ископаемые Для
УДК 564.38:551.782(427+497.11) В. А. Присяжнюк Leiostyla L o v e, 1852 Mollusca, Pulmonata новые ископаемые для неогена Сербии и Украины (Представлено академиком НАН Украины П. Ф. Гожиком) Представители рода Leiostyla Love впервые встречены в сарматских отложениях Сер- бии (в известном местонахождении млекопитающих Врачевич) и в среднем сармате Украины (Волыно-Подолия, Летичевщина). Описаны виды: Leiostyla krstichae из Сербии и Leiostyla ex gr. gottschicki из Украины. В неогене Европы известно лишь несколько видов рода Leiostyla L o v e, результаты изуче- ния которых изложены в работе Дж. Манганелли и др. [1]. Это миоценовые L. gottschicki We n z, описанные В. Венцем из сармата Штейнгейма [2] и Г. Шюттом из риссоевых слоев Австрии [3] и L. austriaca We n z [4] из паннона Австрии; и плиоценовые L. priscilla Pala- dilhe [1], L. capellini S a c c o [1] и L. cf. gottschicki We n z [1]. В последнее время J. K´okay [5] определил молодой экземпляр из сармата Венгрии как L. gottschicki. Этот вид встречен нами в низах нижнего сармата Польши [6]. На мой взгляд, все перечисленные виды, кроме L. austriaca, принадлежат одной группе L. gottschicki We n z. Раковины указанного рода собраны в Центральной Сербии в озерных мергелях и глинах (аналог низов нижнего сармата) в известном местонахождении млекопитающих (Врачевич) и на Подолии (Украина, Летичевщина) в скважине у с. Чапля, также в озерных мергелях средней части среднего сармата. Материал пока хранится в Институте геологических наук НАН Украины в моей коллекции. СЕМЕЙСТВО ORCULIDAE STEENBERG, 1925 Род Leiostyla L o v e, 1852 Leiostyla ex gr. gottschicki We n z, 1921 Табл. -

Conch 40-6-Text.Indb
Journal of ConChology (2011), Vol.40, no.6 583 TAXONOMIC REVISION, HABITATS AND BIOGEOGRAPHY OF THE LAND SNAIL FAMILY DISCIDAE (GASTROPODA: PULMONATA) IN THE CANARY ISLANDS 1 1 2 3 3 DaViD t. holyoak , geralDine a. holyoak , yurena yanes , Maria r. alonso & Miguel iBáñeZ 1Quinta da Cachopa, Barcoila, 6100–014 Cabeçudo, Portugal. 2Instituto Andaluz de Ciencias de la Tierra (CSIC-Universidad de Granada), Camino del Jueves s/n, 18100, Armilla, Granada, Spain. 3Departamento de Biologia Animal, Universidad de La Laguna, E-38206 La Laguna, Tenerife, Islas Canarias, Spain. Abstract The endemic Macaronesian Canaridiscus, provisionally placed in the Discidae genus Atlantica, are closely linked to the peculiar laurel forest habitat of these islands. Knowledge of Atlantica (Canaridiscus) is increased here with the descrip- tion, for the first time, of the genital system of three more of its species. The epiphallus is apparently lacking and the penis is much longer than that known from any of the Discidae of Europe and North America; generally it is too large to be more than partly accommodated in the distal female genital tract. Keraea has been regarded as an endemic Macaronesian genus of Discidae known only from a few shells, but its Madeiran species has been identified as a Trochulus (Hygromiidae) and its type species (from Tenerife) also matches Hygromiidae, not Discidae. The origin and relationships of the Macaronesian Discidae are discussed. The simplest explanatory model is apparently that they represent the last living relicts of a more diverse fauna of Discidae that lived in Europe during the Tertiary; Atlantica (Canaridiscus) was probably isolated early on from the remaining Discidae.