University of Florida Thesis Or Dissertation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Chapter14.Pdf
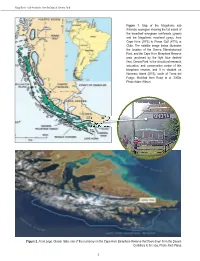
PART I • Omora Park Long-Term Ornithological Research Program THE OMORA PARK LONG-TERM ORNITHOLOGICAL RESEARCH PROGRAM: 1 STUDY SITES AND METHODS RICARDO ROZZI, JAIME E. JIMÉNEZ, FRANCISCA MASSARDO, JUAN CARLOS TORRES-MURA, AND RAJAN RIJAL In January 2000, we initiated a Long-term Ornithological Research Program at Omora Ethnobotanical Park in the world's southernmost forests: the sub-Antarctic forests of the Cape Horn Biosphere Reserve. In this chapter, we first present some key climatic, geographical, and ecological attributes of the Magellanic sub-Antarctic ecoregion compared to subpolar regions of the Northern Hemisphere. We then describe the study sites at Omora Park and other locations on Navarino Island and in the Cape Horn Biosphere Reserve. Finally, we describe the methods, including censuses, and present data for each of the bird species caught in mist nets during the first eleven years (January 2000 to December 2010) of the Omora Park Long-Term Ornithological Research Program. THE MAGELLANIC SUB-ANTARCTIC ECOREGION The contrast between the southwestern end of South America and the subpolar zone of the Northern Hemisphere allows us to more clearly distinguish and appreciate the peculiarities of an ecoregion that until recently remained invisible to the world of science and also for the political administration of Chile. So much so, that this austral region lacked a proper name, and it was generally subsumed under the generic name of Patagonia. For this reason, to distinguish it from Patagonia and from sub-Arctic regions, in the early 2000s we coined the name “Magellanic sub-Antarctic ecoregion” (Rozzi 2002). The Magellanic sub-Antarctic ecoregion extends along the southwestern margin of South America between the Gulf of Penas (47ºS) and Horn Island (56ºS) (Figure 1). -

Wild Patagonia & Central Chile
WILD PATAGONIA & CENTRAL CHILE: PUMAS, PENGUINS, CONDORS & MORE! NOVEMBER 1–18, 2019 Pumas simply rock! This year we enjoyed 9 different cats! Observing the antics of lovely Amber here and her impressive family of four cubs was certainly the highlight in Torres del Paine National Park — Photo: Andrew Whittaker LEADERS: ANDREW WHITTAKER & FERNANDO DIAZ LIST COMPILED BY: ANDREW WHITTAKER VICTOR EMANUEL NATURE TOURS, INC. 2525 WALLINGWOOD DRIVE, SUITE 1003 AUSTIN, TEXAS 78746 WWW.VENTBIRD.COM Sensational, phenomenal, outstanding Chile—no superlatives can ever adequately describe the amazing wildlife spectacles we enjoyed on this year’s tour to this breathtaking and friendly country! Stupendous world-class scenery abounded with a non-stop array of exciting and easy birding, fantastic endemics, and super mega Patagonian specialties. Also, as I promised from day one, everyone fell in love with Chile’s incredible array of large and colorful tapaculos; we enjoyed stellar views of all of the country’s 8 known species. Always enigmatic and confiding, the cute Chucao Tapaculo is in the Top 5 — Photo: Andrew Whittaker However, the icing on the cake of our tour was not birds but our simply amazing Puma encounters. Yet again we had another series of truly fabulous moments, even beating our previous record of 8 Pumas on the last day when I encountered a further 2 young Pumas on our way out of the park, making it an incredible 9 different Pumas! Our Puma sightings take some beating, as they have stood for the last three years at 6, 7, and 8. For sure none of us will ever forget the magical 45 minutes spent observing Amber meeting up with her four 1- year-old cubs as they joyfully greeted her return. -

Wildlife Travel Chile 2018
Chile, species list and trip report, 18 November to 5 December 2018 WILDLIFE TRAVEL v Chile 2018 Chile, species list and trip report, 18 November to 5 December 2018 # DATE LOCATIONS AND NOTES 1 18 November Departure from the UK. 2 19 November Arrival in Santiago and visit to El Yeso Valley. 3 20 November Departure for Robinson Crusoe (Más a Tierra). Explore San Juan Bautista. 4 21 November Juan Fernández National Park - Plazoleta del Yunque. 5 22 November Boat trip to Morro Juanango. Santuario de la Naturaleza Farolela Blanca. 6 23 November San Juan Bautista. Boat to Bahía del Padre. Return to Santiago. 7 24 November Departure for Chiloé. Dalcahue. Parque Tepuhueico. 8 25 November Parque Tepuhueico. 9 26 November Parque Tepuhueico. 10 27 November Dalcahue. Quinchao Island - Achao, Quinchao. 11 28 November Puñihuil - boat trip to Isla Metalqui. Caulin Bay. Ancud. 12 29 November Ferry across Canal de Chacao. Return to Santiago. Farellones. 13 30 November Departure for Easter Island (Rapa Nui). Ahu Tahai. Puna Pau. Ahu Akivi. 14 1 December Anakena. Te Pito Kura. Anu Tongariki. Rano Raraku. Boat trip to Motu Nui. 15 2 December Hanga Roa. Ranu Kau and Orongo. Boat trip to Motu Nui. 16 3 December Hanga Roa. Return to Santiago. 17 4 December Cerro San Cristóbal and Cerro Santa Lucía. Return to UK. Chile, species list and trip report, 18 November to 5 December 2018 LIST OF TRAVELLERS Leader Laurie Jackson West Sussex Guides Claudio Vidal Far South Expeditions Josie Nahoe Haumaka Tours Front - view of the Andes from Quinchao. Chile, species list and trip report, 18 November to 5 December 2018 Days One and Two: 18 - 19 November. -

Movement Behavior, Patch Occupancy, Sustainable Patch Networks and Conservation Planning for an Endemic Understory Bird
MOVEMENT BEHAVIOR, PATCH OCCUPANCY, SUSTAINABLE PATCH NETWORKS AND CONSERVATION PLANNING FOR AN ENDEMIC UNDERSTORY BIRD By TRACI DARNELL CASTELLÓN A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2006 Copyright 2006 by Traci Darnell Castellón ACKNOWLEDGMENTS I sincerely thank my advisor, Kathryn Sieving, for her unwavering support and assistance, and my graduate committee, Lyn Branch, Michael Binford, Graeme Cumming, Doug Levey, and Emelio Bruna. I am also extremely grateful to my field assistants Hector Jañez, John Davis, Alvaro Wurstten, Emma Elgueta, Juan Carlos Correra, computer assistant Nia Haynes, and the many land owners in Chiloé and Osorno who graciously provided access to their farms. This work would not have been possible without their contributions. In addition, I have appreciated and benefited from the support of my friends and colleagues Daniel Smith, Mike Milleson, Matt Reetz, Tom Contreras, Nat Seavy, Marcella Machicote, Greg Jones, and Ivan Díaz. I gratefully acknowledge Mary Willson, Juan Armesto, and Cecilia Smith. Finally, and above all, I thank my parents; Carolyn Blethen and Charles Darnell, my husband Charles Castellón, and my friends, for helping me remember what is important. Partial funding was provided by the Disney Conservation Fund. In-kind support was provided by Fundación Senda Darwin, the University of Florida Map and Image Library, Geoplan Center, the Geography Department, the Land Use and Environmental Change Institute, and the Department of Wildlife Ecology and Conservation. iii TABLE OF CONTENTS page ACKNOWLEDGMENTS ................................................................................................. iii LIST OF TABLES............................................................................................................. vi LIST OF FIGURES ......................................................................................................... -

CARMONA Et Al. (2010) Revista Chilena De Historia Natural 83: 113-142
© Sociedad de Biología de Chile SUPPLEMENTARY MATERIAL CARMONA et al. (2010) Revista Chilena de Historia Natural 83: 113-142. Senda Darwin Biological Station: Long-term ecological research at the interface between science and society Estación Biológica Senda Darwin: Investigación ecológica de largo plazo en la interfase ciencia-sociedad MARTÍN R. CARMONA 1, 2, 5 , J. C. ARAVENA 6, MARCELA A. BUSTAMANTE-SANCHEZ 1, 2 , JUAN L. CELIS-DIEZ 1, 2 , ANDRÉS CHARRIER 2, IVÁN A. DÍAZ 8, JAVIERA DÍAZ-FORESTIER 1, MARÍA F. DÍAZ 1, 10 , AURORA GAXIOLA 1, 2, 5 , ALVARO G. GUTIÉRREZ 7, CLAUDIA HERNANDEZ-PELLICER 1, 3 , SILVINA IPPI 1, 4 , ROCÍO JAÑA-PRADO 1, 2, 9 , PAOLA JARA-ARANCIO 1, 4 , JAIME JIMENEZ 13 , DANIELA MANUSCHEVICH 1, 2 , PABLO NECOCHEA 11 , MARIELA NUÑEZ-AVILA 1, 2, 8 , CLAUDIA PAPIC 11 , CECILIA PÉREZ 2, FERNANDA PÉREZ 1, 2, 5 , SHARON REID 1, 2 , LEONORA ROJAS 1, BEATRIZ SALGADO 1, 2 , CECILIA SMITH- RAMÍREZ 1, 2 , ANDREA TRONCOSO 12 , RODRIGO A. VÁSQUEZ 1, 4 , MARY F. WILLSON 1, RICARDO ROZZI 1 & JUAN J. ARMESTO 1, 2, 5, * 1 Instituto de Ecología y Biodiversidad (IEB), Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Casilla 653, Santiago, Chile 2 Centro de Estudios Avanzados en Ecología y Biodiversidad (CASEB), Departamento de Ecología Pontificia Universidad Católica de Chile, Alameda 340, Casilla 114-D, Santiago, Chile, 833-1150 3 Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Casilla 599 – Raúl Bitrán s/n, Colina El Pino, La Serena, Chile 4 Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Casilla 653, Santiago, Chile 5 Laboratorio Internacional de Cambio Global (LINCGlobal), UC-CSIC, Departamento de Ecología Pontificia Universidad Católica de Chile, Alameda 340, Casilla 114-D, Santiago, Chile, 833-1150 6 Centro de Estudios del Quaternario (CEQUA), Avenida Bulnes 01890, Casilla 737, Punta Arenas, Chile 7 Department of Ecological Modelling, Helmholtz Centre for Environmental Research (UFZ), Permoserstr. -

Chile: North to Patagonia, Nov 2014
Chile: North to Patagonia, Nov 2014 Leader: Fernando Díaz Participants: Cathy Pasterczyk, Marie Carr, Neil Davis, Alain Pataud, Derek Antropik and William Porteous. Manu Expeditions Birding and Wildlife Tours www.Birding-In-Peru.com OVERVIEW…… Chile offers one the most diverse birding experiences in South America. A solid network of domestic flights and an excellent road system make it easy to get around, making it possible to visit a great variety of habitats within just a few days. Mountain passes at 4,500 meters above sea level, arid deserts, scrub, dense forests, steppes and pelagic boat rides on the Pacific Ocean are within easy reach on a journey up and down this long, thin country. The country’s geographic diversity translates into various climates, from arid in the north to Mediterranean in the center and occasionally windy and rainy farther south in Patagonia. This variety is also reflected in the diverse and tasty cuisines that can be enjoyed throughout the journey. Of course, Chile is also world famous for its exquisite wines!. The trip was divided into the three main regions of Chile: North, Central and South/Patagonia. The main goals were to visit as many habitats and to see as many bird species as possible. We included side trips to look for major target species, along with two pelagic boat rides into the rich waters of the Humboldt Current, one from Arica and the other from Valparaiso. Our group of five started in the North, and it was the first time of birding in Chile for most participants. Bill, who had been to Chile’s North on a prior visit, joined us for the Central and Patagonia sections. -

Gear for a Big Year
APPENDIX 1 GEAR FOR A BIG YEAR 40-liter REI Vagabond Tour 40 Two passports Travel Pack Wallet Tumi luggage tag Two notebooks Leica 10x42 Ultravid HD-Plus Two Sharpie pens binoculars Oakley sunglasses Leica 65 mm Televid spotting scope with tripod Fossil watch Leica V-Lux camera Asics GEL-Enduro 7 trail running shoes GoPro Hero3 video camera with selfie stick Four Mountain Hardwear Wicked Lite short-sleeved T-shirts 11” MacBook Air laptop Columbia Sportswear rain shell iPhone 6 (and iPhone 4) with an international phone plan Marmot down jacket iPod nano and headphones Two pairs of ExOfficio field pants SureFire Fury LED flashlight Three pairs of ExOfficio Give- with rechargeable batteries N-Go boxer underwear Green laser pointer Two long-sleeved ExOfficio BugsAway insect-repelling Yalumi LED headlamp shirts with sun protection Sea to Summit silk sleeping bag Two pairs of SmartWool socks liner Two pairs of cotton Balega socks Set of adapter plugs for the world Birding Without Borders_F.indd 264 7/14/17 10:49 AM Gear for a Big Year • 265 Wildy Adventure anti-leech Antimalarial pills socks First-aid kit Two bandanas Assorted toiletries (comb, Plain black baseball cap lip balm, eye drops, toenail clippers, tweezers, toothbrush, REI Campware spoon toothpaste, floss, aspirin, Israeli water-purification tablets Imodium, sunscreen) Birding Without Borders_F.indd 265 7/14/17 10:49 AM APPENDIX 2 BIG YEAR SNAPSHOT New Unique per per % % Country Days Total New Unique Day Day New Unique Antarctica / Falklands 8 54 54 30 7 4 100% 56% Argentina 12 435 -

THE HEART of CHILE January 25-February 8, 2020
® field guides BIRDING TOURS WORLDWIDE [email protected] • 800•728•4953 ITINERARY THE HEART OF CHILE January 25-February 8, 2020 The Inca Tern is one of the most unusual of the tern family. These beautiful birds are found along the Pacific coast of Chile and Peru, and we’ll see them when we visit the coastal areas. Photograph by participant Jane Barnette. We include here information for those interested in the 2020 Field Guides Heart of Chile tour: ¾ a general introduction to the tour ¾ a description of the birding areas to be visited on the tour ¾ an abbreviated daily itinerary with some indication of the nature of each day’s birding outings Those who register for the tour will be sent this additional material: ¾ an annotated list of the birds recorded on a previous year’s Field Guides trip to the area, with comments by guide(s) on notable species or sightings ¾ a detailed information bulletin with important logistical information and answers to questions regarding accommodations, air arrangements, clothing, currency, customs and immigration, documents, health precautions, and personal items ¾ a reference list ¾ a Field Guides checklist for preparing for and keeping track of the birds we see on the tour ¾ after the conclusion of the tour, a list of birds seen on the tour Chile is a country larger than it appears on a map; that long, thin ribbon of land along the west coast of South America is from tip to tip the distance from San Francisco to New York. THE HEART OF CHILE will focus on the central region of the country, allowing for a shorter tour with multiple night stays at most of the hotels we visit. -

Habitat Barriers to Movement of Understory Birds in Fragmented
944 ShortCommunications and Commentaries [Auk, Vol. 113 docarpusNational Park in southern Ecuador(ca. 100 COLLAR, N. J., L. P. GONZAGA, N. KRABBE, A. km north of Angashcola),where recent recordsof S. MADROI•O-NIETO, L. G. NARANJO, T. A. PARKER, cinctusderive (Bloch et al. 1991), and the Reservadel AND D.C. WEGE. 1992. Threatened birds of the Alto Quindio Acaime/Los Nevados National Park/ Americas: The ICBP/IUCN Red Data Book. In- Ucumarl RegionalPark complexin Colombia.Other ternational Council for Bird Preservation, Cam- potentiallyviable populationsof S.cinctus in Ecuador bridge, United Kingdom. probably occurwithin SangayNational Park (Chim- FJELDS•.,J., ANDN. KRABBE.1990. Birdsof the high borazo/Morona-Santiago) and the Cayambe/Coca Andes. Zoological Museum, University of Co- EcologicalReserve (R. S. Ridgely pets. comm.).Strin- penhagen, Denmark. gent protectionof theseareas should be stronglysup- LOISELLE,B. A., AND J. G. BLAKE. 1991. Temporal ported. variation in birds and fruits along an elevational Acknowledgments.--Sponsorsof the Amaluza ex- gradient in CostaRica. Ecology72:180-193. peditions are listed in Williams and Tobias (1994), O'NEILL, J. P., AND T. S. SCHULENBERG.1979. Notes and to these we are indebted. We also thank: Paul on the Masked Saltator, Saltator cinctus, in Peru. Atkinson, ThomasBrooks, Robert Clay, RobertRidge- Auk 96:610-613. ly, and Paul Salaman for reviewing the manuscript RENJIFO,L. M. 1991. Discovery of the Masked Sal- and providing ideas;Rodrigo Tapia-Castro (along with tator in Colombia,with noteson its ecologyand severalmembers of Arcoiris) and David Espinosafor behavior. Wilson Bulletin 103:685-690. assistancein the field; and Nancy Hilgert de Bena- RIDGELY, R. -

Wild Patagonia & Central Chile
WILD PATAGONIA & CENTRAL CHILE: PUMAS, PENGUINS, CONDORS & MORE! October 30 – November 16, 2018 SANTIAGO–HUMBOLDT EXTENSION: ANDES, WETLANDS & ALBATROSS GALORE! November 14-20, 2018 ©2018 Breathtaking Chile! Whether exploring wild Patagonia, watching a Puma hunting a herd of Guanaco against a backdrop of snow-capped spires, enjoying the fascinating antics of a raucous King Penguin colony in Tierra del Fuego, observing a pair of hulking Magellanic Woodpeckers or colorful friendly Tapaculos in a towering Southern Beech forest, or sipping fine wine in a comfortable lodge, this lovely, modern South American country is destined to captivate you! Hosteira Pehoe in Torres Del Paine National Park © Andrew Whittaker Wild Patagonia and Central Chile, Page 2 On this exciting new tour, we will experience the majestic scenery and abundant wildlife of Chile, widely regarded among the most beautiful countries in the world! From Santiago & Talca, in south- central Chile, to the famous Chilean Lake district, charming Chiloe Island to wild Patagonia and Tierra del Fuego in the far south, we will seek out all the special birds, mammals, and vivid landscapes for which the country is justly famous. Our visit is timed for the radiant southern spring when the weather is at its best, colorful blooming wildflowers abound, birds are outfitted in stunning breeding plumage & singing, and photographic opportunities are at their peak. Perhaps most exciting, we will have the opportunity to observe the intimate and poorly known natural history of wild Pumas amid spectacular Torres del Paine National Park, often known as the 8th wonder of the World! Chile is a wonderful place for experiencing nature. -

Adobe PDF, Job 6
Noms français des oiseaux du Monde par la Commission internationale des noms français des oiseaux (CINFO) composée de Pierre DEVILLERS, Henri OUELLET, Édouard BENITO-ESPINAL, Roseline BEUDELS, Roger CRUON, Normand DAVID, Christian ÉRARD, Michel GOSSELIN, Gilles SEUTIN Éd. MultiMondes Inc., Sainte-Foy, Québec & Éd. Chabaud, Bayonne, France, 1993, 1re éd. ISBN 2-87749035-1 & avec le concours de Stéphane POPINET pour les noms anglais, d'après Distribution and Taxonomy of Birds of the World par C. G. SIBLEY & B. L. MONROE Yale University Press, New Haven and London, 1990 ISBN 2-87749035-1 Source : http://perso.club-internet.fr/alfosse/cinfo.htm Nouvelle adresse : http://listoiseauxmonde.multimania. -

(Phthiraptera: Amblycera and Ischnocera) on Birds of Peru
Arxius de Miscel·lània Zoològica, 19 (2021): 7–52 ISSN:Minaya 1698– et0476 al. Checklist of chewing lice (Phthiraptera: Amblycera and Ischnocera) on birds of Peru D. Minaya, F. Príncipe, J. Iannacone Minaya, D., Príncipe, F., Iannacone, J., 2021. Checklist of chewing lice (Phthiraptera: Am- blycera and Ischnocera) on the birds of Peru. Arxius de Miscel·lània Zoològica, 19: 7–52, Doi: https://doi.org/10.32800/amz.2021.19.0007 Abstract Checklist of chewing lice (Phthiraptera: Amblycera and Ischnocera) on birds of Peru. Peru is one of the countries with the highest diversity of birds worldwide, having about 1,876 species in its territory. However, studies focused on chewing lice (Phthiraptera) have been carried out on only a minority of bird species. The available data are distributed in 87 publications in the national and international literature. In this checklist we summarize all the records to date of chewing lice on wild and domestic birds in Peru. Among the 301 species of birds studied, 266 species of chewing lice were recorded. The localities with the highest records were the Departments of Cusco, Junín, Lima and Madre de Dios. No records of birds pa- rasitized by these lice have been found in seven departments of Peru. Studies related to lice have only been reported in 16 % of bird species in the country, indicating that research concerning chewing lice has not yet been performed for the the majority of birds in Peru. Data published through GBIF (Doi: 10.15470/u1jtiu) Key words: Avifauna, Ectoparasites, Lice, Parasitology, Phthiraptera Resumen Lista de verificación de piojos masticadores (Phthiraptera: Amblycera e Ischnocera) de las aves de Perú.