Download (8MB)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Constitution of Kenya Review Commission
CONSTITUTION OF KENYA REVIEW COMMISSION (CKRC) VERBATIM REPORT OF CONSTITUENCY PUBLIC HEARINGS, TINDERET CONSTITUENCY, HELD AT NANDI HILLS TOWN HALL ON 16TH JULY 2002 CONSTITUENCY PUBLIC HEARINGS, TINDERET CONSTITUENCY, HELD AT NANDI HILLS TOWN HALL, ON 16/07/02 Present Com. Alice Yano Com. Isaac Lenaola Apology Com. Prof. Okoth Ogendo Secretariat Staff in Attendance Pauline Nyamweya - Programme Officer Sarah Mureithi - “ Michael Koome - Asst. Prog. Officer Hellen Kanyora - Verbatim Recorder Mr. Barno - District Coordinator 2 The meeting started at 9.30 a.m. with Com. Alice Yano chairing. John Rugut (Chairman 3Cs): Bwana Waziri, the honourable Commissioners, wanakamati wa CCC, mabibi na mabwana nawasalimu tena kwa jina la Yesu hamjambo. Audience: Hatujambo John Rugut: Ningependa kuomba mzee Hezikiah atuongoze kwa sala ili tupate kuanza. (Prayer) Mzee Hezekiah: Tunakushukuru Baba kwa ajili ya mapenzi yako ambayo umetupenda hata ukatujalia siku ya leo ili sisi sote tumefika hapa ili tupate kuyanena na kutazama majadiliano ambayo yako mbele yetu. Kwa hivyo tunaomba msaada kutoka kwako ili nawe utufanyie yaliyo mema ambayo inatatikana kwa mwanadamu ukimuongoza. Asante Baba kwa yote ambayo umetendea na utatutendea mema kwa mengine yote. Hayo tunakushukuru kwa ulinzi wako embayed umetufanyia. Kwa hivi sasa tunakuomba uwe kati yetu tunapoyanena kila jambo lolote tunalosema kutoka katika kinyua chetu. Tunaomba msaada wako, tunaomba uzima wako uwe pamoja nasi sote. Haya ninaweka mikononi mwako katika jina la Yesu aliye muongozi wetu. Amen. Audience: Amen John Rugut: Ningependa wale ambao wamekuwa wakifunza mambo hii ya Katiba (civic educators) wasimame ili Commissioners wapate kuwaona. Hawa ndio civic educators ambao tumekuwa nao katika area hii ya Nandi Hills na kutoka hapa ni George, Bor, Jane, Sally, Joseph na Samuel Rono. -

The Social, Cultural and Economic Impact of Ethnic Violence in Molo Division, 1969 – 2008
THE SOCIAL, CULTURAL AND ECONOMIC IMPACT OF ETHNIC VIOLENCE IN MOLO DIVISION, 1969 – 2008. BY MUIRU PAUL NJOROGE REG. NO. C50/10046/2006 A THESIS SUBMITTED TO THE SCHOOL OF HUMANITIES AND SOCIAL SCIENCES IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE AWARD OF THE DEGREE OF MASTER OF ARTS OF KENYATTA UNIVERSITY. NOVEMBER 2012 ii DECLARATION This thesis is my original work and has not been submitted for a degree in any other university. ______________________Signature Date_______________ Muiru Paul Njoroge Department of History, Archaeology and Political Studies This thesis has been submitted for examination with our knowledge as university supervisors _____________________Signature Date____________________ Dr. Samson Moenga Omwoyo Department of History, Archaeology and Politics Studies ___________________Signature Date____________________ Dr. Peter Wafula Wekesa Department of History, Archaeology and Political Studies iii DEDICATION To my parents, Salome Wanjiru and James Muiru, my brothers and sisters as well was my dear wife, Sharon Chemutai. iv ACKNOWLEDGEMENTS This study would not have been possible without the assistance of several people and institutions. My gratitude first goes to Kenyatta University for awarding me a postgraduate scholarship, and to colleagues at Naivasha Day Secondary School for their moral support. I am eternally indebted to my supervisors Dr Peter Wekesa Wafula and Dr. Samson Moenga Omwoyo who spared no efforts to iniate me into the world of scholarship. I would also like to thank all members of the Department of History, Archaeology and Political Studies for their academic guidance since my undergraduate days (1998-2002). I cannot forget to thank the staff of the Post Modern Library of Kenyatta University, Faculty of Arts and Social Sciences Library of Egerton University for their assistance in the course of writing this work. -

(Still) a Burning Issue: Fuel-Efficient Stove Use in Rural Kenya
(STILL) A BURNING ISSUE: FUEL-EFFICIENT STOVE USE IN RURAL KENYA A Thesis Presented to The Faculty of Graduate Studies of The University of Guelph by ALLISON PATRICIA GRATZ In partial fulfilment of requirements for the degree of Master of Science April, 2009 ©Allison Patricia Gratz, 2009 ABSTRACT (STILL) A BURNING ISSUE: FUEL-EFFICIENT STOVE USE IN RURAL KENYA Allison Patricia Gratz Advisor: University of Guelph, 2009 Professor James P. Mahone Could a stove hold a meaning beyond merely being a tool to cook with? This thesis investigates the knowledge and perceptions of rural residents concerning raw-biomass stoves in Trans Nzoia district, Kenya. The research aimed to identify factors that influenced the use of fuel-efficient stoves in that area. Qualitative data from rural residents, Ministry of Agriculture staff and local administrators were gathered using semi-structured interview techniques and participant observation. Research findings underscored the importance of identifying practical and strategic needs of technology users and the role of local innovation in addressing those needs. There were differences in knowledge and exposure to fuel-efficient stoves depending on the ethnic heterogeneity of an area, age and sex of participants. A conceptual framework developed in the course of this study highlights the importance of culture, exposure, personal factors, and stove characteristics to develop ones capacity to use a new stove. ACKNOWLEDGEMENTS This thesis, although authored by me, would not be in its present form without the help of hundreds of other people. I would like to start by thanking my family, especially my parents, Bob and Verna Duncan. They instilled in me a deep sense of joy and wonderment in discovering the stories and experiences of others quite different from myself, as well as a recognition of the responses that knowledge should bring. -
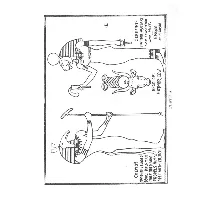
The Origin of the Maasai and Kindred African Tribes and of Bornean Tribes
ct (MEMO' 5(t(I1MET• fiAltOtOHHE4GfO HATHOl\-ASMTAi\f 0E\I1l: HALF MAti AtSO '''fJ1nFHH> \.,/i1'~ NUT. t1At~ Bl~t),WHO HEAD OF· PROPElS HIM:. ENtiAt 5ftFwITI'f ("ufCn MATr4OR, (OW ~F f1AA~AI PLATE: .0'\. PREFACE. The research with which this review deals having been entirely carried out here in Central Africa, far away from all centres of science, the writer is only too well aware that his work must shown signs of the inadequacy of the material for reference at his disposal. He has been obliged to rely entirely on such literature as he could get out from Home, and, in this respect, being obliged for the most psrt to base his selection on the scanty information supplied by publishers' catalogues, he has often had many disappointments when, after months of waiting, the books eventually arrived. That in consequence certain errors may have found their way into the following pages is quite posaible, but he ventures to believe that they are neither many nor of great importance to the subject as a whole. With regard to linguistic comparisons, these have been confined within restricted limits, and the writer has only been able to make comparison with Hebrew, though possibly Aramaic and other Semitic dialects might have carried him further. As there is no Hebrew type in this country he has not been able to give the Hebrew words in their original character as he should have wished. All the quotations from Capt. M. Merker in the following pages are translations of the writer; he is aware that it would have been more correct to have given them in the original German, but in this case they would have been of little value to the majority of the readers of this Journal in Kenya. -

Anthropological Studies in Kavirondo and Nandi Author(S): C
British East Africa: Anthropological Studies in Kavirondo and Nandi Author(s): C. W. Hobley Source: The Journal of the Anthropological Institute of Great Britain and Ireland, Vol. 33 (Jul. - Dec., 1903), pp. 325-359 Published by: Royal Anthropological Institute of Great Britain and Ireland Stable URL: http://www.jstor.org/stable/2842815 . Accessed: 14/06/2014 08:31 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Royal Anthropological Institute of Great Britain and Ireland is collaborating with JSTOR to digitize, preserve and extend access to The Journal of the Anthropological Institute of Great Britain and Ireland. http://www.jstor.org This content downloaded from 91.229.229.101 on Sat, 14 Jun 2014 08:31:36 AM All use subject to JSTOR Terms and Conditions ( 325 ) BRITISH EAST AFRICA. ANTHROPOLOGICAL STUDIES IN KAVIRONDO AND NANDI. BY C. W. HOBLEY, Assoc. M. Ilist. C.E., and AssistantDeputy Commissioner, E.A. Protectorate. [WITH PLATES XXVIII-XXX.] THIS paper contains the results of a more minute researchinto the habits and beliefsof the people already dealt with in a previouspublication of the Institute.' The termEastern Uganda, as applied to this area, is, however,no longercorrect, as since the date of that-publication, the Eastern Province of Uganda has been transferred,and is now known as the Kisumu Province of the East Africa Protectorate. -

Ethnoknowledge of Bukusu Community on Livestock Tick Prevention and Control
Journal of Ethnopharmacology 140 (2012) 298–324 Contents lists available at SciVerse ScienceDirect Journal of Ethnopharmacology jo urnal homepage: www.elsevier.com/locate/jethpharm Ethnoknowledge of Bukusu community on livestock tick prevention and control in Bungoma district, western Kenya a,b,∗ c d e b,f Wycliffe Wanzala , Willem Takken , Wolfgang R. Mukabana , Achola O. Pala , Ahmed Hassanali a School of Pure and Applied Sciences, Department of Biological Sciences, South Eastern University College (A Constituent College of the University of Nairobi), P.O. Box 170-90200, Kitui, Kenya b Behavioural and Chemical Ecology Department (BCED), International Centre of Insect Physiology and Ecology (ICIPE), P.O. Box 30772-00100 GPO, Kasarani, Nairobi, Kenya c Wageningen University and Research Centre, Laboratory of Entomology, P.O. Box 8031, 6700 EH Wageningen, The Netherlands d School of Biological Sciences, University of Nairobi, P.O. Box 30197-00100, Nairobi, Kenya e Technology Transfer Unit, International Centre of Insect Physiology and Ecology (ICIPE), P.O. Box, 30772-00100 GPO, Kasarani, Nairobi, Kenya f School of Pure and Applied Sciences, Kenyatta University, P.O. Box 43844-00100 GPO, Nairobi, Kenya a r t i c l e i n f o a b s t r a c t Article history: Ethnopharmacological relevance: To date, nomadic communities in Africa have been the primary focus Received 24 August 2011 of ethnoveterinary research. The Bukusu of western Kenya have an interesting history, with nomadic Received in revised form lifestyle in the past before settling down to either arable or mixed arable/pastoral farming systems. 14 December 2011 Their collective and accumulative ethnoveterinary knowledge is likely to be just as rich and worth Accepted 13 January 2012 documenting. -

Elixir Journal
49209 Ng’etich Benjamin Kipkios / Elixir Social Studies 113 (2017) 49209-49223 Available online at www.elixirpublishers.com (Elixir International Journal) Social Studies Elixir Social Studies 113 (2017) 49209-49223 Precolonial Nandi Marriage System and Household Relationships Ng‟etich Benjamin Kipkios PhD Candidate, Kisii University, Eldoret Campus, P. O. Box 408, Kisii – 40200, Kenya. ARTICLE INFO Article history: ABSTRACT Received: 19 August 2017; This paper presents the precolonial Nandi marriage system and household relationships Received in revised form: up to the colonial epoch. The study examined the indigenous Nandi family, polygamous 1 December 2017; Nandi household setup, extended family, the clan, incest, and sexual relationships, Accepted: 11 December 2017; responsibilities and rights of household members. It also analyzed the relationships of production, reproduction and distribution, and an analysis of their built forms and Keywords settlement. Precolonial-Nandi, Marriage © 2017 Elixir All rights reserved. System, relationships, Nandi, Clans, households. 1.1 Introduction This paper examines the pre-colonial Nandi marriage system 1.2 Indigenous Nandi Kinship, Marriage, and Religion. and family relationships. This section deals with the 1.2.1 The family. following subtopics; indigenous Nandi Kinship and marriage The English word „family‟ originates from the Latin system, the family, sexual relationships, responsibilities, and familia, meaning „household‟. Lewis Henry Morgan rights of family members. It also covers relations of attempted to trace the origin of the family and came up with a production, reproduction and distribution, access to factors of theory which views with contempt by many anthropologists. production, access to resources and means of biological He argued that there be a close relationship between different reproduction. -

A Collection of 100 Nandi (Kenya) Proverbs and Wise Sayings
A COLLECTION OF 100 NANDI (KENYA) PROVERBS AND WISE SAYINGS BY JOSEPH KAI KARIUKI African Proverbs Working Group NAIROBI KENYA MAY 2017 ACKNOWLEDGEMENT Special thanks go to the African Proverbs Working Group (APWG) Moderator, Father Joseph Healey of the Maryknoll Society for the financial support which helped me to successfully complete this project. I also thank the APWG Committee: Cephas Agbemenu, Margaret Wambere Ireri, Elias Bushiri & Sr. Grace Njau for their support and guidance, not forgetting the APWG Members for their moral support that enabled me to complete this project. Thank you all for your encouragement and support. INTRODUCTION The Location The Nandi are part of the Kalenjin ethnic group, which include eight culturally and linguistically related groups, namely Kipsigis, Tugen, Marakwet, Pokot, Keiyo, Sabaot, Terik and the Nandi. They lived and still form the majority in the highland areas of Nandi Hills in the former Rift Valley Province of Kenya. The Nandi live primarily on high plateaus with good agricultural potential. Ethnographical location of the Nandi of Kenya (Indicated by Red Color) There is a consensus among historians and linguists that plain and highland Nilotes migrated from a region near the southern Ethiopia and Sudan shortly before the beginning of the Christian era and diverged into separate communities shortly thereafter. Kalenjin who already were cattle keepers and had age sets lived in the western Kenya highlands 2,000 years ago. Presumably, these people absorbed other populations already living in the region. From sometime after 500 AD and 1600 AD, there seems to have been a series of migrations eastwards and southwards from near Mount Elgon. -

A History of Kapsabet Girls‟ High School, Nandi County, Kenya, 1960
Asia Pacific Journal of Education, Arts and Sciences, Vol. 3 No. 1, January 2016 ______________________________________________________________________________________________________________ Asia Pacific Journal of A History of Kapsabet Girls‟ High School, Education, Arts and Sciences Nandi County, Kenya, 1960 - 1979 Vol. 3 No.1, 10-20 January 2016 Kiprop Joseph1 & John Koskey Chang’ach2 P-ISSN 2362-8022 Moi University, School of Education, Department of Educational E-ISSN 2362-8030 Foundations www.apjeas.apjmr.com [email protected], [email protected] Date Received: December 1, 2015; Date Revised: December 29, 2015 Abstract - Religious organizations in Kenya are Keywords: Kapsabet, Kenya, Africa Inland empowered by the Education Act to participate in the Mission, Nandi, Africa Inland Church, Oral Interview development and growth of education in schools, (O.I), Local Native Council (LNC), Kenya National colleges and even Universities. Africa Inland Church Archives (KNA) being one of such organizations is not an exception in this case. The paper has endeavoured to establish the INTRODUCTION contribution of Africa Inland Mission/Africa Inland The study area is Nandi Central sub-county. The Church (AIM/AIC) in the development of secondary sub-county is bordered by three sub-counties namely education in Nandi County. The research took Nandi North to the North, Wareng and Nandi East Kapsabet Girls’ High School as a case of its sub-counties to the East and Nandi South and investigation focusing on major events that led to its Kakamega sub-counties to the South and West establishment, growth and the impact of the school to respectively. The sub-county is divided into three the local community. -

Institutions of the Nandi Orkoiyot and Age Set Systems and Their Role in Colonial Resistance in Kenya
International Journal of Science and Research (IJSR) ISSN: 2319-7064 ResearchGate Impact Factor (2018): 0.28 | SJIF (2019): 7.583 Institutions of the Nandi Orkoiyot and Age Set Systems and their Role in Colonial Resistance in Kenya Edwin Kiprop Kemboi1, Samuel A. Nyanchoga2, Melvine C. Lilechi3 1Catholic University of Eastern Africa (CUEA), P. O. Box 62157-00200, Nairobi, Kenya Corresponding author E-mail: edwinkemboi050[at]gmail.com 2Professor of History, Catholic University of Eastern Africa, Nairobi, Kenya E-mail: snyanchoga[at]cuea.edu 3Lecturer, Department of History, Catholic University of Eastern Africa, Nairobi, Kenya E-mail:lilechim[at]cuea.edu Abstract: From the time immemorial, the Nandi community affairs were run through the council of elders in consultation with the seers or prophets. The Nandi community had features of territorial independence and sovereignty as well. However, there is little literature explaining the Nandi traditional security systems before independence. The study attempted to fill the existing gap by investigating the structure and importance of the institution of the Orkoiyot and the age-set system. The study adopted two types of sampling methods, namely the purposive and snowballing. The theoretical framework employed in the study included the structural functionalist theory and social conflict theory. Data collection instruments used in the study included document analysis guide, question guide and focus group discussion guide. The study employed both primary and secondary sources of information. The former was acquired from the archives as well as from oral interviews while the latter was sourced from books and journal articles in several libraries. Since the study was historical, data analysis and presentation was done qualitatively. -

A Gender Analysis of the Influence of Colonial Policies on Access to Land and Agricultural Technology Among the Nandi in Kenya, 1895-1954
International Journal of Humanities and Social Science Vol. 5, No. 9; September 2015 A Gender Analysis of the Influence of Colonial Policies on Access to Land and Agricultural Technology among the Nandi in Kenya, 1895-1954 Prisca J. Tanui Moi University Department Of History Political Science and Public Administration KENYA Abstract This paper examines the agricultural practices in pre-colonial Nandi in relation to the gender question. Specifically, the paper explores and highlights the gender differences in access to agricultural resources and control over them. It examines the historical changes in Nandi’s economic, political and social institutions and demonstrates how these affected production relations. In particular it looks at gender access to land, division of labour, and technology development and how these affected production. The establishment of colonial rule in the region is also examined in this paper. This, in a way, facilitates the analysis of the effects of British conquest on Nandi pre-colonial gender relations in agriculture. The impact of land alienation, forced labour and new technology is examined and so are their influence on gender relations of production and agricultural production in Nandi. The paper is an outcome of a study that used oral data collected in the field, archival material in Kenya National Archives and secondary data, mostly books and journals on the subject from various libraries. The main findings were that political, social and ideological factors interacted in a complex manner and over time influenced gender access to land, control over labour and, technology. Gender relations are essentially perceived as a notion of inequitable power relations. -

Linguistic Stereotypes & Prejudices
Linguistic Stereotypes & Prejudices In Inter-Ethnic Conflicts in Eldoret, Kenya Linguistic Stereotypes & Prejudices In Inter-Ethnic Conflicts in Eldoret, Kenya MARTIN SITUMA Exceller Open Linguistic Stereotypes & Prejudices in Inter-Ethnic Conflicts in Eldoret, Kenya Copyright © Martin Situma, 2020 Martin Situma, Eldoret, Kenya All Rights Reserved Cover design by Exceller Open using resources from Pixabay.com The book has been published with all reasonable efforts taken to make the material error-free after the consent of the author. No portion of the book should be reproduced, stored in any retrieval system (including but not limited to computers, disks, external drives, electronic or digital devices, e-readers, websites), or transmitted in any form or by any means (mechanical, recording, electronic, digital version, photocopying, or otherwise) without the prior, written permission of the publisher, nor be otherwise circulated in any form of binding or cover other than that in which it is published and without a similar condition being imposed on the subsequent purchaser. The views and opinions expressed in this book are the author’s own and the facts are as reported by him. Neither the publisher nor editor endorse or approve the content of this book or guaranty the reliability, accuracy or completeness of the content published herein and do not make and representations of warranties of any kind, express or implied, including but not limited to the implied warranties of merchantability, fitness for a particular purpose. The publisher or editor shall not be liable whatsoever for any errors, omissions, whether such errors or omissions result from negligence, accident or any other cause or claims for loss or damage of any kind, including without limitation damage arising out of use, reliability, accuracy, sufficiency of the information contained in this book.