Marine Science
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

An Analysis of the Afar-Somali Conflict in Ethiopia and Djibouti
Regional Dynamics of Inter-ethnic Conflicts in the Horn of Africa: An Analysis of the Afar-Somali Conflict in Ethiopia and Djibouti DISSERTATION ZUR ERLANGUNG DER GRADES DES DOKTORS DER PHILOSOPHIE DER UNIVERSTÄT HAMBURG VORGELEGT VON YASIN MOHAMMED YASIN from Assab, Ethiopia HAMBURG 2010 ii Regional Dynamics of Inter-ethnic Conflicts in the Horn of Africa: An Analysis of the Afar-Somali Conflict in Ethiopia and Djibouti by Yasin Mohammed Yasin Submitted in partial fulfilment of the requirements for the degree PHILOSOPHIAE DOCTOR (POLITICAL SCIENCE) in the FACULITY OF BUSINESS, ECONOMICS AND SOCIAL SCIENCES at the UNIVERSITY OF HAMBURG Supervisors Prof. Dr. Cord Jakobeit Prof. Dr. Rainer Tetzlaff HAMBURG 15 December 2010 iii Acknowledgments First and foremost, I would like to thank my doctoral fathers Prof. Dr. Cord Jakobeit and Prof. Dr. Rainer Tetzlaff for their critical comments and kindly encouragement that made it possible for me to complete this PhD project. Particularly, Prof. Jakobeit’s invaluable assistance whenever I needed and his academic follow-up enabled me to carry out the work successfully. I therefore ask Prof. Dr. Cord Jakobeit to accept my sincere thanks. I am also grateful to Prof. Dr. Klaus Mummenhoff and the association, Verein zur Förderung äthiopischer Schüler und Studenten e. V., Osnabruck , for the enthusiastic morale and financial support offered to me in my stay in Hamburg as well as during routine travels between Addis and Hamburg. I also owe much to Dr. Wolbert Smidt for his friendly and academic guidance throughout the research and writing of this dissertation. Special thanks are reserved to the Department of Social Sciences at the University of Hamburg and the German Institute for Global and Area Studies (GIGA) that provided me comfortable environment during my research work in Hamburg. -

Zeszyt 10. Morza I Oceany
Uwaga: Niniejsza publikacja została opracowana według stanu na 2008 rok i nie jest aktualizowana. Zamieszczony na stronie internetowej Komisji Standaryzacji Nazw Geograficznych poza Granica- mi Rzeczypospolitej Polskiej plik PDF jest jedynie zapisem cyfrowym wydrukowanej publikacji. Wykaz zalecanych przez Komisję polskich nazw geograficznych świata (Urzędowy wykaz polskich nazw geograficznych świata), wraz z aktualizowaną na bieżąco listą zmian w tym wykazie, zamieszczo- ny jest na stronie internetowej pod adresem: http://ksng.gugik.gov.pl/wpngs.php. KOMISJA STANDARYZACJI NAZW GEOGRAFICZNYCH POZA GRANICAMI RZECZYPOSPOLITEJ POLSKIEJ przy Głównym Geodecie Kraju NAZEWNICTWO GEOGRAFICZNE ŚWIATA Zeszyt 10 Morza i oceany GŁÓWNY URZĄD GEODEZJI I KARTOGRAFII Warszawa 2008 KOMISJA STANDARYZACJI NAZW GEOGRAFICZNYCH POZA GRANICAMI RZECZYPOSPOLITEJ POLSKIEJ przy Głównym Geodecie Kraju Waldemar Rudnicki (przewodniczący), Andrzej Markowski (zastępca przewodniczącego), Maciej Zych (zastępca przewodniczącego), Katarzyna Przyszewska (sekretarz); członkowie: Stanisław Alexandrowicz, Andrzej Czerny, Janusz Danecki, Janusz Gołaski, Romuald Huszcza, Sabina Kacieszczenko, Dariusz Kalisiewicz, Artur Karp, Zbigniew Obidowski, Jerzy Ostrowski, Jarosław Pietrow, Jerzy Pietruszka, Andrzej Pisowicz, Ewa Wolnicz-Pawłowska, Bogusław R. Zagórski Opracowanie Kazimierz Furmańczyk Recenzent Maciej Zych Komitet Redakcyjny Andrzej Czerny, Joanna Januszek, Sabina Kacieszczenko, Dariusz Kalisiewicz, Jerzy Ostrowski, Waldemar Rudnicki, Maciej Zych Redaktor prowadzący Maciej -

Cetacean Rapid Assessment: an Approach to Fill Knowledge Gaps and Target Conservation Across Large Data Deficient Areas
Received: 9 January 2017 Revised: 19 June 2017 Accepted: 17 July 2017 DOI: 10.1002/aqc.2833 RESEARCH ARTICLE Cetacean rapid assessment: An approach to fill knowledge gaps and target conservation across large data deficient areas Gill T. Braulik1,2 | Magreth Kasuga1 | Anja Wittich3 | Jeremy J. Kiszka4 | Jamie MacCaulay2 | Doug Gillespie2 | Jonathan Gordon2 | Said Shaib Said5 | Philip S. Hammond2 1 Wildlife Conservation Society Tanzania Program, Tanzania Abstract 2 Sea Mammal Research Unit, Scottish Oceans 1. Many species and populations of marine megafauna are undergoing substantial declines, while Institute, University of St Andrews, St many are also very poorly understood. Even basic information on species presence is unknown Andrews, Fife, UK for tens of thousands of kilometres of coastline, particularly in the developing world, which is a 3 23 Adamson Terrace, Leven, Fife, UK major hurdle to their conservation. 4 Department of Biological Sciences, Florida 2. Rapid ecological assessment is a valuable tool used to identify and prioritize areas for International University, North Miami, FL, USA conservation; however, this approach has never been clearly applied to marine cetaceans. Here 5 Institute of Marine Science, University of Dar es Salaam, Tanzania a rapid assessment protocol is outlined that will generate broad‐scale, quantitative, baseline Correspondence data on cetacean communities and potential threats, that can be conducted rapidly and cost‐ Gill T. Braulik, Wildlife Conservation Society effectively across whole countries, or regions. Tanzania Program, Zanzibar, Tanzania. Email: [email protected] 3. The rapid assessment was conducted in Tanzania, East Africa, and integrated collection of data on cetaceans from visual, acoustic, and interview surveys with existing information from multiple Funding information sources, to provide low resolution data on cetacean community relative abundance, diversity, and Pew Marine Fellows, Grant/Award Number: threats. -

Investment Opportunities in Africa
A PUBLICATION BY THE AFRICAN AMBASSADORS GROUP IN CAIRO INVESTMENT OPPORTUNITIES IN AFRICA In collaboration with the African Export-Import Bank (Afreximbank) A PUBLICATION BY THE AFRICAN AMBASSADORS GROUP IN CAIRO INVESTMENT OPPORTUNITIES IN AFRICA © Copyright African Ambassadors Group in Cairo, 2018. All rights reserved. African Ambassadors Group in Cairo Email: [email protected] This publication was produced by the African Ambassadors Group in Cairo in collaboration with the African Export-Import Bank (Afreximbank) TABLE OF CONTENTS FOREWORD 8 VOTE OF THANKS 10 INTRODUCTION 12 THE PEOPLE’S DEMOCRATIC REPUBLIC OF ALGERIA 14 THE REPUBLIC OF ANGOLA 18 BURKINA FASO 22 THE REPUBLIC OF BURUNDI 28 THE REPUBLIC OF CAMEROON 32 THE REPUBLIC OF CHAD 36 THE UNION OF COMOROS 40 THE DEMOCRATIC REPUBLIC OF THE CONGO 44 THE REPUBLIC OF CONGO 50 THE REPUBLIC OF CÔTE D’IVOIRE 56 THE REPUBLIC OF DJIBOUTI 60 THE ARAB REPUBLIC OF EGYPT 66 THE STATE OF ERITREA 70 THE FEDERAL DEMOCRATIC REPUBLIC OF ETHIOPIA 74 THE REPUBLIC OF EQUATORIAL GUINEA 78 THE GABONESE REPUBLIC 82 THE REPUBLIC OF GHANA 86 THE REPUBLIC OF GUINEA 90 THE REPUBLIC OF KENYA 94 THE REPUBLIC OF LIBERIA 98 THE REPUBLIC OF MALAWI 102 THE REPUBLIC OF MALI 108 THE REPUBLIC OF MAURITIUS 112 THE KINGDOM OF MOROCCO 116 THE REPUBLIC OF MOZAMBIQUE 120 THE REPUBLIC OF NAMIBIA 126 THE REPUBLIC OF NIGER 130 THE FEDERAL REPUBLIC OF NIGERIA 134 THE REPUBLIC OF RWANDA 138 THE REPUBLIC OF SIERRA LEONE 144 THE FEDERAL REPUBLIC OF SOMALIA 148 THE REPUBLIC OF SOUTH AFRICA 152 THE REPUBLIC OF SOUTH SUDAN 158 THE REPUBLIC OF THE SUDAN 162 THE UNITED REPUBLIC OF TANZANIA 166 THE REPUBLIC OF TUNISIA 170 THE REPUBLIC OF UGANDA 174 THE REPUBLIC OF ZAMBIA 178 THE REPUBLIC OF ZIMBABWE 184 ABOUT AFREXIMBANK 188 FOREWORD Global perception on Africa has positively evolved. -

J.E.Afr.Nat.Hist.Soc. Vol.XXV No.3 (112) January 1966 CHECK LIST of ELOPOID and CLUPEOID FISHES in EAST AFRICAN COASTAL WATERS B
J.E.Afr.Nat.Hist.Soc. Vol.XXV No.3 (112) January 1966 CHECK LIST OF ELOPOID AND CLUPEOID FISHES IN EAST AFRICAN COASTAL WATERS By G.F. LOOSE (East African Marine Fisheries Research Organization, Zanzibar.) Introduction During preliminary biological studies of the economically important fishes of the suborders Elopoidei and Clupeoidei in East African coastal waters, it was found that due to considerable confusion in the existing literature and the taxonomy of many genera, accurate identifi• cations were often difficult. A large collection of elopoid and clupeoid fishes has been made. Specimens have been obtained from purse-seine catches, by trawling in estuaries and shallow bays, by seining, handnetting under lamps at night, and from the catches of indigenous fishermen. A representative fartNaturalof thisHistory),collectionLondon,haswherenow beenI wasdepositedable to examinein the Britishfurther Museummaterial from the Western Indian Ocean during the summer of 1964. Based on these collections this check list has been prepared; a review on the taxonomy, fishery and existing biological knowledge of elopoid and clupeoid species in the East African area is in preparation. Twenty-one species, representing seven families are listed here; four not previously published distributional records are indicated by asterisks. Classification to familial level is based on Whitehead, P.J.P. (1963 a). Keys refer only to species listed and adult fishes. In the synonymy reference is made only to the original description and other subsequent records from the area. Only those localities are listed from which I have examined specimens. East Africa refers to the coastal waters of Kenya and Tanzania and the offshore islands of Pemba, Zanzibar and Mafia; Eastern Africa refers to the eastern side of the African continent, i.e. -

Regional Marine and Coastal Projects in the Western Indian Ocean; an Overview
Participating countries Regional Marine and Coastal Projects in the Western Indian Ocean; an overview WORKING DOCUMENT DATE: 27 May 2009 The purpose of this document is to provide a reference for coastal and marine-related institutions and projects in the Western Indian Ocean region. Content has been included as provided by regional projects or institutions active in more than one country in the Western Indian Ocean. The body of the text describes the activities of 25 projects, but a longer list may be found in Appendix I. Twelve chapters are presented on a thematic or geographic basis, to allow interested projects and parties to interact and coordinate activities around their particular areas of interest. The order of projects is in the order of information received; they are not listed alphabetically or reformatting would have been required after every addition. Sections for which information is pending, or sections that are not applicable have been left blank, also to avoid formatting problems. Much of the text has been extracted from other documents, websites and reports, and no claims are made to originality. This reference document should be used as a guide only. Some information has been based on online sources or project documents that may be outdated. For the most up-to-date, verified and accurate information about any project, please contact them directly. We hope to improve this draft, and so further comments or contributions of any kind are welcome. Compiled by Lucy Scott (ASCLME), with contributions from Tommy Bornman, Juliet -
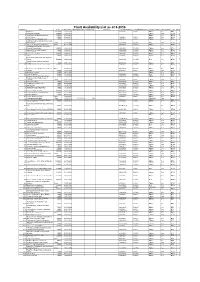
Chart Availability List As of 8-2016
Chart Availability List as of 8-2016 Number Title Scale Edition Date Withdrawn Date Replaced By Replaces Last NM Number Last NM Week-Year Product Status ARCS Chart Folio Disk 2 British Isles 1500000 23.07.2015 325\2016 2-2016 Edition Yes BF6 2 3 Chagos Archipelago 360000 21.06.2012 - Edition Yes BF38 5 5 `Abd Al Kuri to Suqutra (Socotra) 350000 07.03.2013 - Edition Yes BF32 5 6 Gulf of Aden 750000 26.04.2012 124\2015 1-2015 Edition Yes BF32 5 7 Aden Harbour and Approaches 25000 31.10.2013 452\2016 3-2016 Edition Yes BF32 5 La Skhirra-Gabes and Ghannouch with 9 Approaches Plans 24.10.1986 1578\2014 14-2014 New Yes BF24 4 11 Jazireh-Ye Khark and Approaches Plans 03.12.2009 4769\2015 37-2015 Edition Yes BF40 5 Al Aqabah to Duba and Ports on the 12 Coast of Saudi Arabia 350000 14.04.2011 101\2015 53-2015 Edition Yes BF32 5 13 Approaches to Cebu Harbour 35000 21.04.2011 3428\2014 31-2014 Edition Yes BF58 6 14 Cebu Harbour 12500 17.01.2013 4281P\2015 33-2015 Edition Yes BF58 6 15 Approaches to Jizan 200000 17.07.2014 105\2015 53-2015 Edition Yes BF32 5 16 Jizan 30000 19.05.2011 101\2015 53-2015 Edition Yes BF32 5 Plans of the Santa Cruz and Adjacent 17 Islands 500000 14.08.1992 2829\1995 33-1995 New Yes BF68 7 Falmouth Inner Harbour Including 18 Penryn 5000 20.02.2014 5087\2015 40-2015 Edition Yes BF1 1 20 Ile d'Ouessant to Pointe de la Coubre 500000 22.08.2013 419\2016 2-2016 Edition Yes BF16 1 26 Harbours on the South Coast of Devon Plans 17.04.2014 5726\2015 45-2015 Edition Yes BF1 1 27 Bushehr 25000 15.07.2010 984\2016 7-2016 Edition Yes -

Kiszka Et Al Report V2 Rrr Tc
Intended for IOTC-WPEB 2017 use only – Do not cite without consent from the author Cetacean bycatch in the western Indian Ocean: a review of available information on coastal gillnet, tuna purse seine and pelagic longline fisheries Jeremy J. Kiszka1, Per Berggren2, Gill Braulik3, Tim Collins4, Gianna Minton5 & Randall Reeves6 1 Department of Biological Sciences, Florida International University, North Miami, USA 2 School of Natural & Environmental Sciences, Newcastle University, UK 3 Sea Mammal Research Unit, University of St Andrews, United Kingdom. 4 Megaptera Marine Conservation, Wassenaar, the Netherlands 5 Wildlife Conservation Society, Ocean Giants Program, Bronx, New York, USA 6 Okapi Wildlife Associates, Hudson, Quebec, Canada. Email: [email protected] Abstract Bycatch is the most significant and immediate threat to cetacean populations at the global scale. Here we review available information on cetacean bycatch in industrial and small-scale (mostly artisanal) fisheries in the western Indian Ocean (WIO). In coastal waters of the WIO region, the impact of bycatch has not been quantified in small-scale fisheries, except in bottom-set gillnets and driftnets targeting large pelagic fish (including tuna) off Zanzibar. Based on bycatch data from observer programs and estimated abundance of coastal dolphins, the bycatch off the south coast of Zanzibar was found to be unsustainable. Elsewhere in the region, other species are also bycaught, including coastal, oceanic and migratory species such as humpback whales (Megaptera novaeangliae), mostly in bottom-set and drift gillnets. In open-ocean fisheries, bycatch in pelagic longlines has been recorded but seems to be rare. Species affected are mostly medium-sized delphinids (Globicephala macrorhynchus, Grampus griseus, Pseudorca crassidens) involved in depredation (on either bait or catch). -

Physical and Biogeochemical Processes Associated with Upwelling in the Indian Ocean Puthenveettil Narayan
https://doi.org/10.5194/bg-2020-486 Preprint. Discussion started: 9 February 2021 c Author(s) 2021. CC BY 4.0 License. 1 Reviews and syntheses: Physical and biogeochemical processes associated with upwelling in the 2 Indian Ocean 1* 2 3 4 3 Puthenveettil Narayana Menon Vinayachandran , Yukio Masumoto , Mike Roberts , Jenny Hugget , 5 6 7 8 9 4 Issufo Halo , Abhisek Chatterjee , Prakash Amol , Garuda V. M. Gupta , Arvind Singh , Arnab 10 6 11 12 13 5 Mukherjee , Satya Prakash , Lynnath E. Beckley , Eric Jorden Raes , Raleigh Hood 6 7 1Centre for Atmospheric and Oceanic Sciences, Indian Institute of Science, Bengaluru, 560012, India 8 9 2 Graduate School of Science, University of Tokyo, Tokyo, Japan 10 11 3 Nelson Mandela University, Port Elizabeth, South Africa 12 13 4Oceans and Coasts Research, Department of Environment, Forestry and Fisheries, Private Bag X4390, Cape Town 8000, 14 South Africa 15 16 5Department of Conservation and Marine Sciences, Cape Peninsula University of Technology, PO Box 652, Cape Town 17 8000, South Africa 18 6Indian National Centre for Indian Ocean Services, Ministry of Earth Sciences, Hyderabad, India 19 20 7CSIR-National Institute of Oceanography, Regional Centre, Visakhapatnam, 530017, India 21 22 8Centre for Marine Living Resources and Ecology, Ministry of Earth Sciences, Kochi, India 23 24 9Physical Research Laboratory, Ahmedabad, 380009, India 25 26 10 National Centre for Polar and Ocean Research, Ministry of Earth Sciences, Goa, India 27 28 11 Environmental and Conservation Sciences, Murdoch University, Perth, Western Australia 6150, Australia 29 30 12 CSIRO Oceans and Atmosphere, GPO Box 1538, Hobart, TAS, 7001 Australia 31 32 13 University of Maryland Center for Environmental Science, Cambridge, MD, USA 33 34 *Correspondence to: P. -

Annotated Supplement to the Commander's Handbook On
ANNOTATED SUPPLEMENT TO THE COMMANDER’S HANDBOOK ON THE LAW OF NAVAL OPERATIONS NEWPORT, RI 1997 15 NOV 1997 INTRODUCTORY NOTE The Commander’s Handbook on the Law of Naval Operations (NWP 1-14M/MCWP S-2.1/ COMDTPUB P5800.1), formerly NWP 9 (Rev. A)/FMFM l-10, was promulgated to U.S. Navy, U.S. Marine Corps, and U.S. Coast Guard activities in October 1995. The Com- mander’s Handbook contains no reference to sources of authority for statements of relevant law. This approach was deliberately taken for ease of reading by its intended audience-the operational commander and his staff. This Annotated Supplement to the Handbook has been prepared by the Oceans Law and Policy Department, Center for Naval Warfare Studies, Naval War College to support the academic and research programs within the College. Although prepared with the assistance of cognizant offices of the General Counsel of the Department of Defense, the Judge Advocate General of the Navy, The Judge Advocate General of the Army, The Judge Advocate General of the Air Force, the Staff Judge Advo- cate to the Commandant of the Marine Corps, the Chief Counsel of the Coast Guard, the Chairman, Joint Chiefs of Staff and the Unified Combatant Commands, the annotations in this Annotated Supplement are not to be construed as representing official policy or positions of the Department of the Navy or the U.S. Governrnent. The text of the Commander’s Handbook is set forth verbatim. Annotations appear as footnotes numbered consecutively within each Chapter. Supplementary Annexes, Figures and Tables are prefixed by the letter “A” and incorporated into each Chapter. -

A Viability Assessment of Using Bath Sponge and Pearl Oyster Farms to Filter Highly Olluted Waters in the Zanzibar Channel Hayley Oakland SIT Study Abroad
SIT Graduate Institute/SIT Study Abroad SIT Digital Collections Independent Study Project (ISP) Collection SIT Study Abroad Spring 2013 Bioremediation Mariculture in Zanzibar, Tanzania: A Viability Assessment of Using Bath Sponge and Pearl Oyster Farms to Filter Highly olluted Waters in the Zanzibar Channel Hayley Oakland SIT Study Abroad Follow this and additional works at: https://digitalcollections.sit.edu/isp_collection Part of the Aquaculture and Fisheries Commons, Environmental Health and Protection Commons, Environmental Indicators and Impact Assessment Commons, Marine Biology Commons, and the Natural Resources and Conservation Commons Recommended Citation Oakland, Hayley, "Bioremediation Mariculture in Zanzibar, Tanzania: A Viability Assessment of Using Bath Sponge and Pearl Oyster Farms to Filter Highly olluted Waters in the Zanzibar Channel" (2013). Independent Study Project (ISP) Collection. 1526. https://digitalcollections.sit.edu/isp_collection/1526 This Unpublished Paper is brought to you for free and open access by the SIT Study Abroad at SIT Digital Collections. It has been accepted for inclusion in Independent Study Project (ISP) Collection by an authorized administrator of SIT Digital Collections. For more information, please contact [email protected]. Bioremediation Mariculture in Zanzibar, Tanzania: A viability assessment of using bath sponge and pearl oyster farms to filter highly polluted waters in the Zanzibar Channel Hayley Oakland SIT Spring 2013 Zanzibar: Coastal Ecology and Natural Resource Management University of Oregon, Department of Marine Biology and Environmental Science Advisor: Dr. Aviti J. Mmochi Academic Advisor: Nat Quansah Table of Contents Acknowledgements 1 Abstract 2 I. Introduction 3 II. Study Area 11 III. Methods 12 3.1 Biodiversity and Farming Surveys 12 a. -

Tanzania Biodiversity Threats Assessment
Tanzania Biodiversity Threats Assessment Biodiversity threats and management opportunities for SUCCESS in Fumba, Bagamoyo, and Mkuranga This publication is available electronically on the Coastal Resources Center’s website: www.crc.uri.edu. It is also available on the Western Indian Ocean Marine Science Organization’s website: www.wiomsa.org. For more information contact: Coastal Resources Center, University of Rhode Island, Narragansett Bay Campus, South Ferry Road, Narragansett, RI 02882, USA. Email: [email protected] Citation: Torell, Elin, Mwanahija Shalli, Julius Francis, Baraka Kalangahe, Renalda Munubi, 2007, Tanzania Biodiversity Threats Assessment: Biodiversity Threats and Management Opportunities for Fumba, Bagamoyo, and Mkuranga, Coastal Resources Center, University of Rhode Island, Narragansett, 47 pp. Disclaimer: This report was made possible by the generous support of the American people through the United States Agency for International Development (USAID). The contents are the responsibility of the authors and do not necessarily reflect the views of USAID or the United States Government. Cooperative agreement # EPP-A-00-04-00014-00 Cover Photo: Beach scene from Bagamoyo Photo Credit: Elin Torell EXECUTIVE SUMMARY The Sustainable Coastal Communities and Ecosystems (SUCCESS) Program falls under the Congressional biodiversity earmark, where it fits under the secondary code. These are programs and activities – site based or not – that have biodiversity conservation as an explicit, but not primary objective. One criterion for such programs is that their activities must be defined based on an analysis of threats to biodiversity. This report aims to assess the biodiversity threats in the land-seascapes where SUCCESS operates. The purpose is to understand the major direct threats to biodiversity as well as the context and root causes of the threats.