Sinopsis De La Familia Acanthaceae En El Perú
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pollen Diversity Studies in Some Taxa of Bicarpellatae from Nagpur
Pollen Diversity Studies in Some Taxa of Bicarpellatae from Nagpur Sapna V. Awachat Department of Botany, Rashtrasant Tukadoji Maharaj Nagpur University, Nagpur Abstract: Dicots are the diverse group of plants on the basis of morphology, anatomy, cytogenetics, embryology and pollen morphology. Pollen diversity is the study of the variations in the morphology of pollen grains. Pollen has also proved to be an excellent tool in taxonomic studies. The application of pollen characters in solving controversial taxonomical and phylogenetic problems has now been widely recognized all over the world (Mandal, 2010). In the present paper, pollen diversity of several taxa belonging to Bicarpellatae (Apocynaceae, Asclepiadaceae, Boraginaceae, Convolvulaceae, Solanaceae, Scrophulariaceae, Acanthaceae, Thunbergiaceae, Verbenaceae, Lamiaceae, etc.) from Nagpur is selected. The pollen grains show variations with respect to exine ornamentation, aperture type, shape, size and NPC (number, position, character) classification etc. The pollen slides were prepared by using acetolysis method (Erdtman 1952) and documentation was done by using light microscope and digital camera. The pollen grain studies show variation in exine ornamentation (psilate to verrucate), aperture (porate to spiraperturate), shape (oblate to prolate) and size (small to large). The pollens described on the basis of NPC classification are presented. Pollen calendar and distribution was also noted in all taxa. It is found that the pollen grains with relative variations in pollen morphology help us to differentiate families. Keywords : Bicarpellatae, Pollen grains, Acetolysis. Introduction: Palynology involves the study of pollen and encompasses the structural and functional aspect of pollen. Pollen grains come in an infinite variety of shapes with complex surface ornamentation and occur on almost every surface in nature. -

Modelando La Distribución De Atlantea Tulita : Especie Endémica En Peligro
ModelandoModelando lala distribucidistribuci óónn dede AtlanteaAtlantea tulitatulita :: especieespecie endend éémicamica enen peligropeligro dede extinciextinci óónn enen PuertoPuerto RicoRico Andr és V élez Estudiante de Maestr ía Universidad de Puerto Rico, Mayag üez Departamento de Biolog ía Franz Lab - Insect Systematics & Evolution http:// academic.uprm.edu /franz /ave.html IntroducciIntroducci óónn yy justificacijustificaci óónn • Perdida de diversidad Modificación paisaje • Endemismos Cambio climático (Thomas et al 2004) • Condición de Puerto Rico Pérdida de habitad • Conocimiento para CONSERVAR • Herramienta Modelación potencial distribución • Áreas protegidas conocer la actual o futura distribución EstatusEstatus A.A. tulitatulita -Lista 88 especies amenazadas DRNA (Departamento de Recursos Naturales y Ambientales de Puerto Rico) 2007 (Protección federal) -El DRNA (2009) 1°invertebrado lista de especies en peligro de extinción -Petición comunidad (2010) por destrucción de hábitat -Presencia restringida a el municipio Quebradillas y otra en el municipio de Maricao (Carrión, 2003) -Especie en amenaza? -Falta de investigaciones para concluir • Elaborar un modelo de predicción de distribución • Área idóneas en Puerto Rico • Trabajos de bilogía y ecología • Pautas para conservación ObjetivoObjetivo Realizar un modelo de predicción de la distribución potencial para la mariposa endémica A. tulita , actualmente en peligro de extinción en Puerto Rico ÁÁrearea dede estudioestudio PuertoPuerto RicoRico -Las-Área temperaturas aproximada dey lluvias 8990 km2 • El norte fresco y lluvioso -temperaturas Isla principal deentre Puerto 13-32°C Rico, •Mona, El sur semiáridoVieques y temperaturasCulebra. entre 13-38°C •-Diversidad El centro montañoso de ecosistemas: lluvioso y frío,0-1330m. promedio Bosques lluvias secos, 2540 mm temperaturasB. lluviosos, zona entre cársica, 4-32°C. áreas montañosas, -ecosistemas La cobertura costeros vegetal sey ha desarrolladomarinos. -

Indigenous Use and Ethnopharmacology of Medicinal Plants in Far-West Nepal
Indigenous Use and Ethnopharmacology of Medicinal Plants in Far-west Nepal Ripu M. Kunwar, Y. Uprety, C. Burlakoti, C.L. Chowdhary and R.W. Bussmann Research Abstract Ethnopharmacological knowledge is common and im- jor part of these therapies. Interest in phytomedicine is port among tribal populations but much of the information also renewed during the last decade and many medicinal is empirical at best lacking scientific validation. Despite plant species are being screened for pharmacological ac- widespread use of plant resources in traditional medi- tivities. The global demand of herbal medicine is growing cines, bioassay analysis of very few plant species have and its market is expanding at the rate of 20% annually in been conducted to investigate their medicinal properties, India (Srivastava 2000, Subrat 2002). The world market and to ascertain safety and efficacy of traditional reme- for herbal remedies in 1999 was worth of U.S.$19.4 bil- dies. The present study analyses indigenous uses of me- lion (Laird & Pierce 2002). dicinal plants of far-west Nepal and compares with earlier ayurveda studies, phytochemical assessments and phar- Numerous drugs have entered into the international mar- macological actions. A field study was carried out in Baita- ket through exploration of ethnopharmacology and tradi- di and Darchula districts of far-west Nepal. Group discus- tional medicine (Bussmann 2002) with extensive uses of sions, informal meetings, questionnaire surveys and field medicinal plants. It is estimated that 25% of prescription observations were employed for primary data collection. drugs contain active principles derived from higher plants (Tiwari & Joshi 1990). The first compound derived from Voucher specimens were collected with field notes and herbal remedies to enter the international market was codes and deposited at Tribhuvan University Central Her- ephedrine, an amphetamine like stimulant from Ephed- barium (TUCH), Kathmandu. -

Acanthaceae), a New Chinese Endemic Genus Segregated from Justicia (Acanthaceae)
Plant Diversity xxx (2016) 1e10 Contents lists available at ScienceDirect Plant Diversity journal homepage: http://www.keaipublishing.com/en/journals/plant-diversity/ http://journal.kib.ac.cn Wuacanthus (Acanthaceae), a new Chinese endemic genus segregated from Justicia (Acanthaceae) * Yunfei Deng a, , Chunming Gao b, Nianhe Xia a, Hua Peng c a Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou, 510650, People's Republic of China b Shandong Provincial Engineering and Technology Research Center for Wild Plant Resources Development and Application of Yellow River Delta, Facultyof Life Science, Binzhou University, Binzhou, 256603, Shandong, People's Republic of China c Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming, 650201, People's Republic of China article info abstract Article history: A new genus, Wuacanthus Y.F. Deng, N.H. Xia & H. Peng (Acanthaceae), is described from the Hengduan Received 30 September 2016 Mountains, China. Wuacanthus is based on Wuacanthus microdontus (W.W.Sm.) Y.F. Deng, N.H. Xia & H. Received in revised form Peng, originally published in Justicia and then moved to Mananthes. The new genus is characterized by its 25 November 2016 shrub habit, strongly 2-lipped corolla, the 2-lobed upper lip, 3-lobed lower lip, 2 stamens, bithecous Accepted 25 November 2016 anthers, parallel thecae with two spurs at the base, 2 ovules in each locule, and the 4-seeded capsule. Available online xxx Phylogenetic analyses show that the new genus belongs to the Pseuderanthemum lineage in tribe Justi- cieae. -

Hypoestes Aristata (Vahl) Sol
Biol Res 43: 403-409, 2010 BHATT ET AL. Biol Res 43, 2010, 403-409 B403R The foliar trichomes of Hypoestes aristata (Vahl) Sol. ex Roem. & Schult var aristata (Acanthaceae) a widespread medicinal plant species in tropical sub-Saharan Africa: with comments on its possible phylogenetic significance A. Bhatt*, Y. Naidoo and A. Nicholas School of Biological and Conservation Sciences, University of KwaZulu-Natal, Westville Campus, Private Bag X54001, Durban, KZN, 4000, South Africa ABSTRACT The micromorphology of foliar trichomes of Hypoestes aristata var. aristata was studied using stereo, light and scanning microscopy (SEM). This genus belongs to the advanced angiosperm family Acanthaceae, for which few micromorphological leaf studies exist. Results revealed both glandular and non-glandular trichomes, the latter being more abundant on leaf veins, particularly on the abaxial surface of very young leaves. With leaf maturity, the density of non-glandular trichomes decreased. Glandular trichomes were rare and of two types: long-stalked capitate and globose-like peltate trichomes. Capitate trichomes were observed only on the abaxial leaf surface, while peltate trichomes were distributed on both adaxial and abaxial leaf surfaces. Key terms: Acanthaceae, Glandular trichomes, Hypoestes aristata var. aristata, medicinal plant, Scanning electron microscope. INTRODUCTION zygomorphic flowers supported by prominent bracts and producing explosive capsular fruits. Many studies have The Family Acanthaceae is a large and diverse family of further supported the placement of Hypoestes in a smaller dicotyledonous plants comprising about 202 genera and 3520 clade that includes the prominent genus Justicia (McDade species (Judd et al., 2008); although estimates vary from 2600 and Moody 1999). -
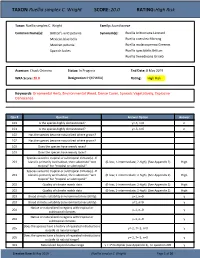
TAXON:Ruellia Simplex C. Wright SCORE:20.0 RATING:High Risk
TAXON: Ruellia simplex C. Wright SCORE: 20.0 RATING: High Risk Taxon: Ruellia simplex C. Wright Family: Acanthaceae Common Name(s): Britton's wild petunia Synonym(s): Ruellia brittoniana Leonard Mexican blue bells Ruellia coerulea Morong Mexican petunia Ruellia malacosperma Greenm. Spanish ladies Ruellia spectabilis Britton Ruellia tweedieana Griseb. Assessor: Chuck Chimera Status: In Progress End Date: 8 May 2019 WRA Score: 20.0 Designation: H(HPWRA) Rating: High Risk Keywords: Ornamental Herb, Environmental Weed, Dense Cover, Spreads Vegetatively, Explosive Dehiscence Qsn # Question Answer Option Answer 101 Is the species highly domesticated? y=-3, n=0 n 101 Is the species highly domesticated? y=-3, n=0 n 102 Has the species become naturalized where grown? 102 Has the species become naturalized where grown? 103 Does the species have weedy races? 103 Does the species have weedy races? Species suited to tropical or subtropical climate(s) - If 201 island is primarily wet habitat, then substitute "wet (0-low; 1-intermediate; 2-high) (See Appendix 2) High tropical" for "tropical or subtropical" Species suited to tropical or subtropical climate(s) - If 201 island is primarily wet habitat, then substitute "wet (0-low; 1-intermediate; 2-high) (See Appendix 2) High tropical" for "tropical or subtropical" 202 Quality of climate match data (0-low; 1-intermediate; 2-high) (See Appendix 2) High 202 Quality of climate match data (0-low; 1-intermediate; 2-high) (See Appendix 2) High 203 Broad climate suitability (environmental versatility) y=1, n=0 -

The Bear in Eurasian Plant Names
Kolosova et al. Journal of Ethnobiology and Ethnomedicine (2017) 13:14 DOI 10.1186/s13002-016-0132-9 REVIEW Open Access The bear in Eurasian plant names: motivations and models Valeria Kolosova1*, Ingvar Svanberg2, Raivo Kalle3, Lisa Strecker4,Ayşe Mine Gençler Özkan5, Andrea Pieroni6, Kevin Cianfaglione7, Zsolt Molnár8, Nora Papp9, Łukasz Łuczaj10, Dessislava Dimitrova11, Daiva Šeškauskaitė12, Jonathan Roper13, Avni Hajdari14 and Renata Sõukand3 Abstract Ethnolinguistic studies are important for understanding an ethnic group’s ideas on the world, expressed in its language. Comparing corresponding aspects of such knowledge might help clarify problems of origin for certain concepts and words, e.g. whether they form common heritage, have an independent origin, are borrowings, or calques. The current study was conducted on the material in Slavonic, Baltic, Germanic, Romance, Finno-Ugrian, Turkic and Albanian languages. The bear was chosen as being a large, dangerous animal, important in traditional culture, whose name is widely reflected in folk plant names. The phytonyms for comparison were mostly obtained from dictionaries and other publications, and supplemented with data from databases, the co-authors’ field data, and archival sources (dialect and folklore materials). More than 1200 phytonym use records (combinations of a local name and a meaning) for 364 plant and fungal taxa were recorded to help find out the reasoning behind bear-nomination in various languages, as well as differences and similarities between the patterns among them. Among the most common taxa with bear-related phytonyms were Arctostaphylos uva-ursi (L.) Spreng., Heracleum sphondylium L., Acanthus mollis L., and Allium ursinum L., with Latin loan translation contributing a high proportion of the phytonyms. -

Acanthaceae and Asteraceae Family Plants Used by Folk Medicinal Practitioners for Treatment of Malaria in Chittagong and Sylhet Divisions of Bangladesh
146 American-Eurasian Journal of Sustainable Agriculture, 6(3): 146-152, 2012 ISSN 1995-0748 ORIGINAL ARTICLE Acanthaceae and Asteraceae family plants used by folk medicinal practitioners for treatment of malaria in Chittagong and Sylhet Divisions of Bangladesh Md. Tabibul Islam, Protiva Rani Das, Mohammad Humayun Kabir, Shakila Akter, Zubaida Khatun, Md. Megbahul Haque, Md. Saiful Islam Roney, Rownak Jahan, Mohammed Rahmatullah Faculty of Life Sciences, University of Development Alternative, Dhanmondi, Dhaka-1205, Bangladesh Md. Tabibul Islam, Protiva Rani Das, Mohammad Humayun Kabir, Shakila Akter, Zubaida Khatun, Md. Megbahul Haque, Md. Saiful Islam Roney, Rownak Jahan, Mohammed Rahmatullah: Acanthaceae and Asteraceae family plants used by folk medicinal practitioners for treatment of malaria in Chittagong and Sylhet Divisions of Bangladesh ABSTRACT Malaria is a debilitating disease causing high mortality rates among men and women if not treated properly. The disease is prevalent in many countries of the world with the most prevalence noted among the sub-Saharan countries, where it is in an epidemic form. The disease is classified as hypo-endemic in Bangladesh with the southeast and the northeastern regions of the country having the most malaria-affected people. The rural people suffer most from malaria, and they rely on folk medicinal practitioners for treatment, who administer various plant species for treatment of the disease as well as associated symptoms like pain and fever. Plant species have always formed the richest sources of anti-malarial drugs, the most notable being quinine and artemisinin. However, quinine has developed drug-resistant vectors and artemisinin is considered by some to developing initial resistance, particularly in China, where it has been used for thousands of years to combat malaria. -

Landscape Plant List
APPENDIX B-Tree Technical Manual, Download at the "Unified Development Code" from: http://www.cityofedinburg.com/ City of Edinburg Native (Permitted) Plant List e e = P Wildlif s t rac espan: Scientific Name Family Common Name(s) Slow) Medium, Fast, COMMENTS Perennial, A=Annual, D=deciduous Period Blooming Color Bloom Aquatic Soils Moist Riparian Upland Full Shade Shade/Sun Full Sun Att Lif (Bi=Bird Bu=Butterfly(Bi=Bird Be=Bee Height Mature Width Mature Rate Growth ( Spacing Large Trees (Parking lot shade) Acacia wrightii Fabaceae Wright's Acacia X X X Be 30' 20' Medium 20' P, D Spring White Recurved spines; heat & drought tolerant Fast growing shade tree; small fruit is extremely valuable for birds; limbs fairly Celtis laevigata Ulmaceae Sugar Hackberry X X X X X Bi 45' 50' Fast 50' P, D Spring Greenish brittle; drops fine, sticky sap, which is messy Fragrant, showy clusters of small, white flowers produce large quantities of fruit Ehretia anacua Boraginaceae Anacua X X X Bi 45' 50' Slow 50' P, D Jun-Oct White valuable to wildlife; fruit drop can be messy; good shade tree Large, spreading tree that requires regular watering to reach full potential; Fraxinus berlandieriana Oleaceae Mexican Ash, Fresno X X X X Bi 50' 75' Medium 75' P, D Spring Greenish papery, winged fruits on female trees only Very fast growing tree, but relatively Tepeguaje, Lead Leucaena pulverulenta Fabaceae X X Be 40' 50' Fast 50' P, D Spring Summer White short lived; limbs brittle and break easily, Tree and subject to girdling beetles Dense shade tree provides important -

Atoll Research Bulletin No. 503 the Vascular Plants Of
ATOLL RESEARCH BULLETIN NO. 503 THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS BY NANCY VANDER VELDE ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C., U.S.A. AUGUST 2003 Uliga Figure 1. Majuro Atoll THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS ABSTRACT Majuro Atoll has been a center of activity for the Marshall Islands since 1944 and is now the major population center and port of entry for the country. Previous to the accompanying study, no thorough documentation has been made of the vascular plants of Majuro Atoll. There were only reports that were either part of much larger discussions on the entire Micronesian region or the Marshall Islands as a whole, and were of a very limited scope. Previous reports by Fosberg, Sachet & Oliver (1979, 1982, 1987) presented only 115 vascular plants on Majuro Atoll. In this study, 563 vascular plants have been recorded on Majuro. INTRODUCTION The accompanying report presents a complete flora of Majuro Atoll, which has never been done before. It includes a listing of all species, notation as to origin (i.e. indigenous, aboriginal introduction, recent introduction), as well as the original range of each. The major synonyms are also listed. For almost all, English common names are presented. Marshallese names are given, where these were found, and spelled according to the current spelling system, aside from limitations in diacritic markings. A brief notation of location is given for many of the species. The entire list of 563 plants is provided to give the people a means of gaining a better understanding of the nature of the plants of Majuro Atoll. -

UNIVERSIDAD DE GUADALAJARA COORDINACIÓN GENERAL ACADÉMICA Coordinación De Bibliotecas Biblioteca Digital
UNIVERSIDAD DE GUADALAJARA COORDINACIÓN GENERAL ACADÉMICA Coordinación de Bibliotecas Biblioteca Digital La presente tesis es publicada a texto completo en virtud de que el autor ha dado su autorización por escrito para la incorporación del documento a la Biblioteca Digital y al Repositorio Institucional de la Universidad de Guadalajara, esto sin sufrir menoscabo sobre sus derechos como autor de la obra y los usos que posteriormente quiera darle a la misma. Av. Hidalgo 935, Colonia Centro, C.P. 44100, Guadalajara, Jalisco, México [email protected] - Tel. 31 34 22 77 ext. 11959 UNIVERSIDAD DE GUADALAJARA Centro Universitario de los Lagos Maestría en Ciencia y Tecnología “Evaluación del efecto ansiolítico del extracto metanólico de hojas de Justicia spicigera (Schltdl.) y sus fracciones en ratas hembra en las fases de proestro- estro”. TESIS PARA OBTENER EL GRADO DE MAESTRO EN CIENCIA Y TECNOLOGÍA CON ORIENTACIÓN EN BIOMÉDICA PRESENTA: IBQ. ANA RAQUEL RAMOS MOLINA Director de tesis: Dr. en C. Cesar Soria Fregozo Codirector de tesis: Dr. en C. Armando Mora Pérez Lagos de Moreno, Jal., junio de 2019. II El presente trabajo se realizó en el laboratorio de Ciencias Biomédicas (Área Histología y Psicobiología) y Síntesis de Química Orgánica del Centro Universitario de los Lagos de la Universidad de Guadalajara, bajo la Dirección del Dr. Cesar Soria Fregozo y la Co-Dirección del Dr. Armando Mora Pérez. El proyecto recibió apoyo financiero parcial del PROGRAMA DE APOYO A LA MEJORA EN LAS CONDICIONES DE PRODUCCIÓN DE LOS MIEMBROS DEL SNI Y SNCA (PRO-SNI) convocatoria 2017 y 2018. Durante la realización del presente trabajo Ana Raquel Ramos Molina recibió una beca del Consejo Nacional de Ciencia y Tecnología (CONACyT) para estudios de posgrado con número de registro CVU 855016. -

Downloaded and Set As out Groups Genes
Alzahrani et al. BMC Genomics (2020) 21:393 https://doi.org/10.1186/s12864-020-06798-2 RESEARCH ARTICLE Open Access Complete chloroplast genome sequence of Barleria prionitis, comparative chloroplast genomics and phylogenetic relationships among Acanthoideae Dhafer A. Alzahrani1, Samaila S. Yaradua1,2*, Enas J. Albokhari1,3 and Abidina Abba1 Abstract Background: The plastome of medicinal and endangered species in Kingdom of Saudi Arabia, Barleria prionitis was sequenced. The plastome was compared with that of seven Acanthoideae species in order to describe the plastome, spot the microsatellite, assess the dissimilarities within the sampled plastomes and to infer their phylogenetic relationships. Results: The plastome of B. prionitis was 152,217 bp in length with Guanine-Cytosine and Adenine-Thymine content of 38.3 and 61.7% respectively. It is circular and quadripartite in structure and constitute of a large single copy (LSC, 83, 772 bp), small single copy (SSC, 17, 803 bp) and a pair of inverted repeat (IRa and IRb 25, 321 bp each). 131 genes were identified in the plastome out of which 113 are unique and 18 were repeated in IR region. The genome consists of 4 rRNA, 30 tRNA and 80 protein-coding genes. The analysis of long repeat showed all types of repeats were present in the plastome and palindromic has the highest frequency. A total number of 98 SSR were also identified of which mostly were mononucleotide Adenine-Thymine and are located at the non coding regions. Comparative genomic analysis among the plastomes revealed that the pair of the inverted repeat is more conserved than the single copy region.