Mandibular Fossa of Homo Erectus
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -
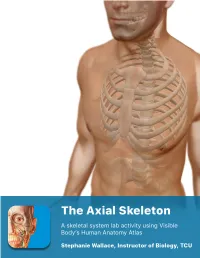
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

MBB: Head & Neck Anatomy
MBB: Head & Neck Anatomy Skull Osteology • This is a comprehensive guide of all the skull features you must know by the practical exam. • Many of these structures will be presented multiple times during upcoming labs. • This PowerPoint Handout is the resource you will use during lab when you have access to skulls. Mind, Brain & Behavior 2021 Osteology of the Skull Slide Title Slide Number Slide Title Slide Number Ethmoid Slide 3 Paranasal Sinuses Slide 19 Vomer, Nasal Bone, and Inferior Turbinate (Concha) Slide4 Paranasal Sinus Imaging Slide 20 Lacrimal and Palatine Bones Slide 5 Paranasal Sinus Imaging (Sagittal Section) Slide 21 Zygomatic Bone Slide 6 Skull Sutures Slide 22 Frontal Bone Slide 7 Foramen RevieW Slide 23 Mandible Slide 8 Skull Subdivisions Slide 24 Maxilla Slide 9 Sphenoid Bone Slide 10 Skull Subdivisions: Viscerocranium Slide 25 Temporal Bone Slide 11 Skull Subdivisions: Neurocranium Slide 26 Temporal Bone (Continued) Slide 12 Cranial Base: Cranial Fossae Slide 27 Temporal Bone (Middle Ear Cavity and Facial Canal) Slide 13 Skull Development: Intramembranous vs Endochondral Slide 28 Occipital Bone Slide 14 Ossification Structures/Spaces Formed by More Than One Bone Slide 15 Intramembranous Ossification: Fontanelles Slide 29 Structures/Apertures Formed by More Than One Bone Slide 16 Intramembranous Ossification: Craniosynostosis Slide 30 Nasal Septum Slide 17 Endochondral Ossification Slide 31 Infratemporal Fossa & Pterygopalatine Fossa Slide 18 Achondroplasia and Skull Growth Slide 32 Ethmoid • Cribriform plate/foramina -

Traumatic Dislocation of the Mandibular Condyle Into the Middle Cranial Fossa Treated with Immediate Reconstruction: a Case Report Joseph E
CILLO, SINN, AND ELLIS 859 grafting in a patient with AIDS, acute myocardial infarction, fection and acquired immunodeficiency syndrome. Am J Surg and severe left main coronary artery disease. J Cardiovasc Surg 180:228, 2000 44:55, 2003 18. James J, Hofland HW, Borgstein ES, et al: The prevalence of HIV 15. van Marle J, Tudhope L, Weir G, et al: Vascular disease in infection among burn patients in a burns unit in Malawi and its HIV/AIDS patients. S Afr Med J 92:974, 2002 influence on outcome. Burns 29:55, 2003 16. Chambers AJ, Lord RS: Incidence of acquired immune defi- 19. Patton LL, Shugars DA, Bonito AJ: A systematic review of ciency syndrome (AIDS)-related disorders at laparatomy in pa- complication risks for HIV-positive patients undergoing inva- tients with AIDS. Br J Surg 88:294, 2001 sive dental procedures. J Am Dent Assoc 133:195, 2002 17. Tran HS, Moncure M, Tarnoff M, et al: Predictors of operative 20. Diz Dios P, Fernandez Feijoo J, Vazquez Garcia E: Tooth ex- outcome in patients with human immunodeficiency virus in- traction in HIV sero-positive patients. Int Dent J 49:317, 1999 J Oral Maxillofac Surg 63:859-865, 2005 Traumatic Dislocation of the Mandibular Condyle into the Middle Cranial Fossa Treated With Immediate Reconstruction: A Case Report Joseph E. Cillo, Jr, DMD,* Douglas P. Sinn, DDS,† and Edward Ellis III, DDS, MS‡ While fractures of the mandible condylar process are his chief complaints were right-sided jaw pain, limited common, accounting for 29% to 40% of mandibular mouth opening, and malocclusion with an anterior open trauma,1 dislocation of the mandibular condyle through bite. -

Inferior View of Skull
human anatomy 2016 lecture sixth Dr meethak ali ahmed neurosurgeon Inferior View Of Skull the anterior part of this aspect of skull is seen to be formed by the hard palate.The palatal process of the maxilla and horizontal plate of the palatine bones can be identified . in the midline anteriorly is the incisive fossa & foramen . posterolaterlly are greater & lesser palatine foramena. Above the posterior edge of the hard palate are the choanae(posterior nasal apertures ) . these are separated from each other by the posterior margin of the vomer & bounded laterally by the medial pterygoid plate of sphenoid bone . the inferior end of the medial pterygoid plate is prolonged as a curved spike of bone , the pterygoid hamulus. the superior end widens to form the scaphoid fossa . posterolateral to the lateral pterygoid plate the greater wing of the sphenoid is pieced by the large foramen ovale & small foramen spinosum . posterolateral to the foramen spinosum is spine of the sphenoid . Above the medial border of the scaphoid fossa , the sphenoid bone is pierced by pterygoid canal . Behind the spine of the sphenoid , in the interval between the greater wing of the sphenoid and the petrous part of the temporal bone , there is agroove for the cartilaginous part of the auditory tube. The opening of the bony part of the tube can be identified. The mandibular fossa of the temporal bone & the articular tubercle form the upper articular surfaces for the temporomandibular joint . separating the mandibular fossa from the tympanic plate posteriorly is the squamotympanic fissure, through the medial end of which (petrotympanic fissure ) the chorda tympani exits from the tympanic cavity .The styloid process of the temporal bone projects downward & forward from its inferior aspect. -

Skull / Cranium
Important! 1. Memorizing these pages only does not guarantee the succesfull passing of the midterm test or the semifinal exam. 2. The handout has not been supervised, and I can not guarantee, that these pages are absolutely free from mistakes. If you find any, please, report to me! SKULL / CRANIUM BONES OF THE NEUROCRANIUM (7) Occipital bone (1) Sphenoid bone (1) Temporal bone (2) Frontal bone (1) Parietal bone (2) BONES OF THE VISCEROCRANIUM (15) Ethmoid bone (1) Maxilla (2) Mandible (1) Zygomatic bone (2) Nasal bone (2) Lacrimal bone (2) Inferior nasalis concha (2) Vomer (1) Palatine bone (2) Compiled by: Dr. Czigner Andrea 1 FRONTAL BONE MAIN PARTS: FRONTAL SQUAMA ORBITAL PARTS NASAL PART FRONTAL SQUAMA Parietal margin Sphenoid margin Supraorbital margin External surface Frontal tubercle Temporal surface Superciliary arch Zygomatic process Glabella Supraorbital margin Frontal notch Supraorbital foramen Internal surface Frontal crest Sulcus for superior sagittal sinus Foramen caecum ORBITAL PARTS Ethmoidal notch Cerebral surface impresiones digitatae Orbital surface Fossa for lacrimal gland Trochlear notch / fovea Anterior ethmoidal foramen Posterior ethmoidal foramen NASAL PART nasal spine nasal margin frontal sinus Compiled by: Dr. Czigner Andrea 2 SPHENOID BONE MAIN PARTS: CORPUS / BODY GREATER WINGS LESSER WINGS PTERYGOID PROCESSES CORPUS / BODY Sphenoid sinus Septum of sphenoid sinus Sphenoidal crest Sphenoidal concha Apertura sinus sphenoidalis / Opening of sphenoid sinus Sella turcica Hypophyseal fossa Dorsum sellae Posterior clinoid process Praechiasmatic sulcus Carotid sulcus GREATER WINGS Cerebral surface • Foramen rotundum • Framen ovale • Foramen spinosum Temporal surface Infratemporalis crest Infratemporal surface Orbital surface Maxillary surface LESSER WINGS Anterior clinoid process Superior orbital fissure Optic canal PTERYGOID PROCESSES Lateral plate Medial plate Pterygoid hamulus Pterygoid fossa Pterygoid sulcus Scaphoid fossa Pterygoid notch Pterygoid canal (Vidian canal) Compiled by: Dr. -

Osteoplastic Orbitozygomatic-Mastoid- Transattical Craniotomy Craniotomia Orbitozigomático-Mastóideo-Transatical Osteoplástica
THIEME Technical Note | Nota Técnica 257 Osteoplastic Orbitozygomatic-Mastoid- Transattical Craniotomy Craniotomia orbitozigomático-mastóideo-transatical osteoplástica Ilton Guenhiti Shinzato1,2,3,4 Felipe Bouchabki de Almeida Guardini4,5 Fernando Kobayashi4,5 1 Neurosurgery Residency Program Supervisor of Santa Casa, Universidade Address for correspondence Ilton Guenhiti Shinzato, MD, DSc, Federal do Mato Grosso do Sul, Campo Grande, MS, Brazil Neurosurgery Service, Santa Casa Hospital, Rua Eduardo Santos 2 DSc, COPPE/UFRJ, Rio de Janeiro, RJ, Brazil; Thesis Research done at Pereira, 88, 79002-251, Campo Grande, MS, Brazil Teikyo University, Chiba, Japan (e-mail: [email protected]). 3 TitularMemberofABNc,ABORL,FLANC,WFNS;ActiveMemberof the Walter E. Dandy Neurosurgical Society, St. Louis, United States 4 Titular Member of Brazilian Neurosurgery Society – SBN, São Paulo, SP, Brazil 5 Neurosurgeon, Residency at Santa Casa, Universidade Federal do Mato Grosso do Sul, Campo Grande, MS, Brazil Arq Bras Neurocir 2016;35:257–269. Abstract Objective The objective of this study is to introduce and describe a surgical technique called Osteoplastic Orbitozygomatic-Mastoid-Transattical Craniotomy (Osteoplastic OZ-MT) and to show the possibility of accomplishing osteoplastic craniotomies for other classical lateral transcranial approaches. Technique The Osteoplastic OZ-MT combines many lateral transcranial approaches. The surgical approach involves structures of lateral and basal portions of the skull, from the frontal bone, superolateral-inferolateral-posterolateral walls of the orbit, zygoma, zygomatic process, sphenoid greater and lesser wings, temporal fossa, mandibular fossa, zygomatic process of the temporal bone, petrous pyramid, mastoid, up to the parietal and occipital regions. The temporal muscle is totally preserved and attached to the one-piece-only bone flap. -

Skeletal System + Joints)
Chapters 610 (Skeletal system + joints) Exercise 6: Introduction to the Skeletal System. Figure 6.1 Axial or Appendicular (Divisions) Figure 6.2 Major bones in the body Figure 6.3 Bone “Classification” Figure 6.5‐6.6 Bone model and slides (parts) Table 6.2 Terminology Exercise 7: Appendicular Skeleton Scapula: o Acromion process o Scapular notch o Coracoid process o Supraspinous fossa o Glenoid Cavity o Spine o Lateral (axillary) border o Subscapular fossa o Inferior angle o Medial boarder o Superior angle o Infraspinous fossa o Superior boarder Clavicle o Coronoid turbercle o Sternal end o Acromial end Humerous o Head o Olecranon fossa o Greater turbercle o Lateral epicondyle o Lesser tubercle o Medial epicondyle o Deltoid tuberosity o Capitulum o Radial fossa o Trochlea o Coronoid fossa Radius o Head o Ulnar notch o Neck o Styloid process o Radial tuberosity Ulna o Olecranon process o Radial notch o Trochlear notch o Head of the ulna o Coronoid process o Styloid process Hand o Carpals o Metacarpals o Phalanges Pelvic girdle (7.6 and 7.7) o Sacrum o Coxal bone o Sacrolilliac joint . Illium o Acetabulum . Pubis o Symphysis pubis . Ischium o Obturator foramen o Ischial tuberosity o Iliac crest o Ischial ramus o Illiac fossa Femur o Head o Fovea capitis o Greater trochanter o Body o Neck o Lateral epicondyle o Intertrochanteric line o Medial epicondyle (anterior) o Patellar groove o Intertrochanteric crest o Medial condyle (posterior) o Lateral condyle o Lesser trochanter o Intercondylar fossa Patella o Base o Apex Tibia -

Surgical Anatomy of the Temporal Bone Gülay Açar and Aynur Emine Çiçekcibaşı
Chapter Surgical Anatomy of the Temporal Bone Gülay Açar and Aynur Emine Çiçekcibaşı Abstract Numerous neurological lesions and tumors of the paranasal sinuses and oral cavity may spread into the middle and posterior cranial fossae through the ana- tomical apertures. For the appropriate management of these pathologies, many extensive surgical approaches with a comprehensive overview of the anatomical landmarks are required from the maxillofacial surgery’s point of view. The surgical significance lies in the fact that iatrogenic injury to the petrous segment of the tem- poral bone including the carotid artery, sigmoid sinus, and internal jugular vein, can lead to surgical morbidity and postoperative pseudoaneurysm, vasospasm, or carotid-cavernous fistula. To simplify understanding complex anatomy of the temporal bone, we aimed to review the surgical anatomy of the temporal bone focusing on the associations between the surface landmarks and inner structures. Also, breaking down an intricate bony structure into smaller parts by compart- mental approach could ease a deep concentration and navigation. To identify the anatomic architecture of the temporal bone by using reference points, lines and compartments can be used to supplement anatomy knowledge of maxillofacial surgeons and may improve confidence by surgical trainees. Especially, this system- atic method may provide an easier way to teach and learn surgical spatial structure of the petrous pyramid in clinical applications. Keywords: maxillofacial surgery, segmentation, surface landmarks, surgical anatomy, temporal bone 1. Introduction The temporal bone is a dense complex bone that constitutes the lower lateral aspect of the skull and has complex anatomy because of the three-dimensional relationships between neurovascular structures. -

Temporomandibular Joint (TMJ)
Regions of the Head 9. Oral Cavity & Perioral Regions Temporomandibular Joint (TMJ) Zygomatic process of temporal bone Articular tubercle Mandibular Petrotympanic fossa of TMJ fissure Postglenoid tubercle Styloid process External acoustic meatus Mastoid process (auditory canal) Atlanto- occipital joint Fig. 9.27 Mandibular fossa of the TMJ tubercle. Unlike other articular surfaces, the mandibular fossa is cov- Inferior view of skull base. The head (condyle) of the mandible artic- ered by fi brocartilage, not hyaline cartilage. As a result, it is not as ulates with the mandibular fossa of the temporal bone via an articu- clearly delineated on the skull (compare to the atlanto-occipital joints). lar disk. The mandibular fossa is a depression in the squamous part of The external auditory canal lies just posterior to the mandibular fossa. the temporal bone, bounded by an articular tubercle and a postglenoid Trauma to the mandible may damage the auditory canal. Head (condyle) of mandible Pterygoid Joint fovea capsule Neck of Coronoid mandible Lateral process ligament Neck of mandible Lingula Mandibular Stylomandibular foramen ligament Mylohyoid groove AB Fig. 9.28 Head of the mandible in the TMJ Fig. 9.29 Ligaments of the lateral TMJ A Anterior view. B Posterior view. The head (condyle) of the mandible is Left lateral view. The TMJ is surrounded by a relatively lax capsule that markedly smaller than the mandibular fossa and has a cylindrical shape. permits physiological dislocation during jaw opening. The joint is stabi- Both factors increase the mobility of the mandibular head, allowing lized by three ligaments: lateral, stylomandibular, and sphenomandib- rotational movements about a vertical axis. -

The Skeleton AXIAL SKELETON
BIOL 231, Human Anatomy Wennstrom The Skeleton Goals: • Identify the bones and bone features listed below • List the articulations of each of the bones below and identify the sites of articulation (e.g. which bones make joints with which other bones, and which bone parts form those joints?) • Distinguish whether a specimen is from the left or right side of the body for the scapula, humerus, coxal bone, and femur AXIAL SKELETON CRANIAL BONES FACIAL BONES VERTEBRAL COLUMN Frontal bone Nasal bones Atlas coronal suture Maxilla transverse foramen supraorbital foramen incisive foramen Axis orbital surface palatine process transverse foramen Parietal bones median palatine suture dens lambdoidal suture infraorbital foramen Cervical vertebrae (7) sagittal suture orbital surface Thoracic vertebrae (12) squamosal suture Lacrimal bones Lumbar vertebrae (5) Occipital bone Zygomatic bones Sacrum (5 fused bones) foramen magnum temporal process Coccyx (4 fused bones) occipital condyles orbital surface Vertebral structures Temporal bones Palatine bones centrum (body) external acoustic meatus horizontal plate vertebral foramen internal acoustic meatus Vomer superior & inferior articular styloid process Mandible facets zygomatic process angle vertebral arch (lamina & mastoid process body pedicle) mandibular fossa mandibular foramen transverse process carotid canal mental foramen spinous process jugular foramen mandibular notch stylomastoid foramen mental protuberance THORAX Sphenoid bone alveolar margin Rib sella -

An Investigation of the Morphology of the Petrotympanic Fissure Using Cone-Beam Computed Tomography
JOURNAL OF ORAL & MAXILLOFACIAL RESEARCH Damaskos et al. An Investigation of the Morphology of the Petrotympanic Fissure Using Cone-Beam Computed Tomography Spyros Damaskos1,2, Konstantinos Syriopoulos3,4, Rogier L. Sens3,4, Constantinus Politis3,4 1Department of Oral Radiology, Academic Centre for Dentistry Amsterdam, Amsterdam, The Netherlands. 2Department of Oral Diagnosis and Radiology, School of Dentistry, NKUA, Athens, Greece. 3Department of Imaging and Pathology, OMFS IMPATH Research Group, Faculty of Medicine, Katholieke Universiteit Leuven, Leuven, Belgium. 4Department of Oral and Maxillofacial Surgery, University Hospitals Leuven, Leuven, Belgium. Corresponding Author: Spyros Damaskos Department of Oral Radiology Academic Centre for Dentistry Amsterdam (ACTA) Gustav Mahler Laan 3004, 1081 LA, Amsterdam The Netherlands Phone: +30 6944 541529 E-mail: [email protected]; [email protected] ABSTRACT Objectives: The purpose of the present study was: a) to examine the visibility and morphology of the petrotympanic fissure on cone-beam computed tomography images, and b) to investigate whether the petrotympanic fissure morphology is significantly affected by gender and age, or not. Material and Methods: Using Newtom VGi (QR Verona, Italy), 106 cone-beam computed tomography examinations (212 temporomandibular joint areas) of both genders were retrospectively and randomly selected. Two observers examined the images and subsequently classified by consensus the petrotympanic fissure morphology into the following three types: type 1 - widely open; type 2 - narrow middle; type 3 - very narrow/closed. Results: The petrotympanic fissure morphology was assessed as type 1, type 2, and type 3 in 85 (40.1%), 72 (34.0%), and 55 (25.9%) cases, respectively. No significant difference was found between left and right petrotympanic fissure morphology (Kappa = 0.37; P < 0.001).