Title PHYLOGENETIC SPECULATION of the TUNICATA Author
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Review Sheet for Lecture 15: Life in the Benthic Realm
Lecture 15: Life in the Benthic Realm Terminology Planktonic, nektonic, benthic, epifaunal, infaunal, sessile, mobile, bathyal, abyssal, abyssopelagic, hadopelagic, supratidal, intertidal, subtidal, spicules, spongin, radial symmetry, encruster, epidermal cell, porocyte, sclerocyte, amoebocyte, mesohyl, choanocyte, polyp, medusa, colonial, solitary, aragonite, octocoral, gorgonin, cnidocyst, nematocyst, zooxanthellae, hematypic, coral bleaching, lophophore, zoid, zooecium, zooaria, polymorphism, pedicle, helically coiled shell, radially coiled shell, exoskeleton, endoskeleton Dates: none Review Questions: Be familiar with the life and feeding modes of all of the invertebrate groups we discussed and their major morphologies, behaviors, life strategies etc. Why are so many sessile invertebrates radially symmetric? Explain the three different wall types found in sponges, what’s the point? Discuss the different roles of the different sponge cells How do hematypic corals contribute to the creation of different nearshore environments and conditions? Describe the community created by an innkeeper worm How does the skeleton of a brachiopod differ from that of a clam? How do the shells of clams, snails, and nautiloids differ? Classification: Phylum Porifera Class Demospongiae (soft sponges) Class Calcarea (calcareous sponges) Class Hexactinellida (glass sponges) Phylum Cnidaria Class Anthozoa -stony corals -sea fans -soft corals -sea anemones -sea pens Phylum Annelida (worms) -bristleworms -innkeeper worm -lugworms -spaghetti worm -feather duster -catworm Phylum Bryozoa (moss-animals) Phylum Mollusca -Class Bivalvia (clams) -Class Gastropoda (snails and slugs) -Class Cephalopoda (squid, octopus, nautiloid) -Class Scaphopoda (tusk shells) -Class Polyplacophora (chitons) . -

Invertebrates Invertebrates: • Are Animals Without Backbones • Represent 95% of the Animal Kingdom Animal Diversity Morphological Vs
Invertebrates Invertebrates: • Are animals without backbones • Represent 95% of the animal kingdom Animal Diversity Morphological vs. Molecular Character Phylogeny? A tree is a hypothesis supported or not supported by evidence. Groupings change as new evidence become available. Sponges - Porifera Natural Bath Sponges – over-collected, now uncommon Sponges • Perhaps oldest animal phylum (Ctenphora possibly older) • may represent several old phyla, some now extinct ----------------Ctenophora? Sponges - Porifera • Mostly marine • Sessile animals • Lack true tissues; • Have only a few cell types, cells kind of independent • Most have no symmetry • Body resembles a sac perforated with holes, system of canals. • Strengthened by fibers of spongin, spicules Sponges have a variety of shapes Sponges Pores Choanocyte Amoebocyte (feeding cell) Skeletal Water fiber flow Central cavity Flagella Choanocyte in contact with an amoebocyte Sponges - Porifera • Sessile filter feeder • No mouth • Sac-like body, perforated by pores. • Interior lined by flagellated cells (choanocytes). Flagellated collar cells generate a current, draw water through the walls of the sponge where food is collected. • Amoeboid cells move around in the mesophyll and distribute food. Sponges - Porifera Grantia x.s. Sponge Reproduction Asexual reproduction • Fragmentation or by budding. • Sponges are capable of regeneration, growth of a whole from a small part. Sexual reproduction • Hermaphrodites, produce both eggs and sperm • Eggs and sperm released into the central cavity • Produces -

S I Section 4
3/31/2011 Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Porifera Ecdysozoa Deuterostomia Lophotrochozoa Cnidaria and Ctenophora Cnidaria and Protostomia SSiection 4 Radiata Bilateria Professor Donald McFarlane Parazoa Eumetazoa Lecture 13 Invertebrates: Parazoa, Radiata, and Lophotrochozoa Ancestral colonial choanoflagellate 2 Traditional classification based on Parazoa – Phylum Porifera body plans Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Parazoa 4 main morphological and developmental Sponges features used Loosely organized and lack Porifera Ecdysozoa Cnidaria and Ctenophora tissues euterostomia photrochozoa D 1. o Presence or absence of different tissue L Multicellular with several types of types Protostomia cells 2. Type of body symmetry Radiata Bilateria 8,000 species, mostly marine Parazoa Eumetazoa 3. Presence or absence of a true body No apparent symmetry cavity Ancestral colonial Adults sessile, larvae free- choanoflagellate 4. Patterns of embryonic development swimming 3 4 1 3/31/2011 Water drawn through pores (ostia) into spongocoel Flows out through osculum Reproduce Choanocytes line spongocoel Sexually Most hermapppgggphrodites producing eggs and sperm Trap and eat small particles and plankton Gametes are derived from amoebocytes or Mesohyl between choanocytes and choanocytes epithelial cells Asexually Amoebocytes absorb food from choanocytes, Small fragment or bud may detach and form a new digest it, and carry -

Practical Paleoecology GEO 342
Practical Paleoecology GEO 342 Compiled By: Dr. Osama Attia Compiled By Dr. Osama Attia 1 Phylum Brachiopoda Brachiopods or lamp shells Name: Name means "arm" (brachio) + "foot" (pod). Main characteristics: - Bivalved (two shells), each with bilateral symmetry. - The plane of symmetry passes through the center of each shell or valve. - The two valves differ in size and shape in most. Sometimes the larger valve will have an opening near the hinge line through which the pedicle extended in life. - Soft parts include a lophophore consisting of coiled tentacles with cilia. The lophophore circulates water between the two valves, distributing oxygen and flushing out carbon dioxide. Water movements caused by the lophophore also transport food particles toward the mouth. Geologic range: Lower Cambrian to Recent. Very abundant during the Paleozoic. A few species (belonging to only three families) remain today. Mode of life: Inhabitants of shallow marine environments; they generally live attached in a fixed position on the seafloor. Inarticulate brachiopods are known to live in burrows in the sediment. Brachiopods are filter feeders. Compiled By Dr. Osama Attia 2 Living positions of brachiopods. A = Articulate brachiopod attached to the seafloor by its pedicle. B = Interior of brachiopod valve showing lophophore. C = Inarticulate brachiopod, Lingula, which lives within a tube or burrow in seafloor sediment. A. CLASS INARTICULATA – The Inarticulate Brachiopods Primitive brachiopods with phosphatic or chitinous valves; no hinge. Spoon-shaped valves held together with muscles and soft parts. Lingula is a well known inarticulate brachiopod. Geologic range: Lower Cambrian to Recent Compiled By Dr. Osama Attia 3 Fossil inarticulate brachiopod from the Cambrian. -

Animal Diversity Part 2
Textbook resources • pp. 517-522 • pp. 527-8 Animal Diversity • p. 530 part 2 • pp. 531-2 Clicker question In protostomes A. The blastopore becomes the mouth. B. The blastopore becomes the anus. C. Development involves indeterminate cleavage. D. B and C Fig. 25.2 Phylogeny to know (1). Symmetry Critical innovations to insert: Oral bilateral symmetry ecdysis mouth develops after anus multicellularity Aboral tissues 1 Animal diversity, part 2 Parazoa Diversity 2 I. Parazoa • Porifera: Sponges II. Cnidaria & Ctenophora • Tissues • Symmetry I. Outline the • Germ Layers III. Lophotrochozoa unique • Embryonic characteristics Development of sponges IV. Ecdysozoa • Body Cavities • Segmentation Parazoa Parazoa • Porifera: Sponges • Porifera: Sponges – Multicellular without – Hermaphrodites tissues – Sexual and asexual reproduction – Choanocytes (collar cells) use flagella to move water and nutrients into pores – Intracellular digestion Fig. 25.11 Animal diversity, part 2 Clicker Question Diversity 2 I. Parazoa In diploblastic animals, the inner lining of the digestive cavity or tract is derived from II. Cnidaria & Ctenophora A. Endoderm. II. Outline the B. Ectoderm. unique III. Lophotrochozoa C. Mesoderm. characteristics D. Coelom. of cnidarians and IV. Ecdysozoa ctenophores 2 Coral Box jelly Cnidaria and Ctenophora • Cnidarians – Coral; sea anemone; jellyfish; hydra; box jellies • Ctenophores – Comb jellies Sea anemone Jellyfish Hydra Comb jelly Cnidaria and Ctenophora Fig. 25.12 Coral Box jelly Cnidaria and Ctenophora • Tissues Fig. 25.12 – -

Tunicata 4 Alberto Stolfi and Federico D
Tunicata 4 Alberto Stolfi and Federico D. Brown Chapter vignette artwork by Brigitte Baldrian. © Brigitte Baldrian and Andreas Wanninger. A. Stolfi Department of Biology , Center for Developmental Genetics, New York University , New York , NY , USA F. D. Brown (*) EvoDevo Laboratory, Departamento de Zoologia , Instituto de Biociências, Universidade de São Paulo , São Paulo , SP , Brazil Evolutionary Developmental Biology Laboratory, Department of Biological Sciences , Universidad de los Andes , Bogotá , Colombia Centro Nacional de Acuicultura e Investigaciones Marinas (CENAIM) , Escuela Superior Politécnica del Litoral (ESPOL) , San Pedro , Santa Elena , Ecuador e-mail: [email protected] A. Wanninger (ed.), Evolutionary Developmental Biology of Invertebrates 6: Deuterostomia 135 DOI 10.1007/978-3-7091-1856-6_4, © Springer-Verlag Wien 2015 [email protected] 136 A. Stolfi and F.D. Brown Above all , perhaps , I am indebted to a decidedly the phylogenetic relationships between the three vegetative , often beautiful , and generally obscure classes and many orders and families have yet to group of marine animals , both for their intrinsic interest and for the enjoyment I have had in search- be satisfactorily settled. Appendicularia, ing for them . N. J. Berrill (1955) Thaliacea, and Ascidiacea remain broadly used in textbooks and scientifi c literature as the three classes of tunicates; however, recent molecular INTRODUCTION phylogenies have provided support for the mono- phyly of only Appendicularia and Thaliacea, but Tunicates are a group of marine fi lter-feeding not of Ascidiacea (Swalla et al. 2000 ; animals1 that have been traditionally divided into Tsagkogeorga et al. 2009 ; Wada 1998 ). A para- three classes: (1) Appendicularia, also known as phyletic Ascidiacea calls for a reevaluation of larvaceans because their free-swimming and tunicate relationships. -

Nemertean and Phoronid Genomes Reveal Lophotrochozoan Evolution and the Origin of Bilaterian Heads
Nemertean and phoronid genomes reveal lophotrochozoan evolution and the origin of bilaterian heads Author Yi-Jyun Luo, Miyuki Kanda, Ryo Koyanagi, Kanako Hisata, Tadashi Akiyama, Hirotaka Sakamoto, Tatsuya Sakamoto, Noriyuki Satoh journal or Nature Ecology & Evolution publication title volume 2 page range 141-151 year 2017-12-04 Publisher Springer Nature Macmillan Publishers Limited Rights (C) 2017 Macmillan Publishers Limited, part of Springer Nature. Author's flag publisher URL http://id.nii.ac.jp/1394/00000281/ doi: info:doi/10.1038/s41559-017-0389-y Creative Commons Attribution 4.0 International (http://creativecommons.org/licenses/by/4.0/) ARTICLES https://doi.org/10.1038/s41559-017-0389-y Nemertean and phoronid genomes reveal lophotrochozoan evolution and the origin of bilaterian heads Yi-Jyun Luo 1,4*, Miyuki Kanda2, Ryo Koyanagi2, Kanako Hisata1, Tadashi Akiyama3, Hirotaka Sakamoto3, Tatsuya Sakamoto3 and Noriyuki Satoh 1* Nemerteans (ribbon worms) and phoronids (horseshoe worms) are closely related lophotrochozoans—a group of animals including leeches, snails and other invertebrates. Lophotrochozoans represent a superphylum that is crucial to our understand- ing of bilaterian evolution. However, given the inconsistency of molecular and morphological data for these groups, their ori- gins have been unclear. Here, we present draft genomes of the nemertean Notospermus geniculatus and the phoronid Phoronis australis, together with transcriptomes along the adult bodies. Our genome-based phylogenetic analyses place Nemertea sis- ter to the group containing Phoronida and Brachiopoda. We show that lophotrochozoans share many gene families with deu- terostomes, suggesting that these two groups retain a core bilaterian gene repertoire that ecdysozoans (for example, flies and nematodes) and platyzoans (for example, flatworms and rotifers) do not. -
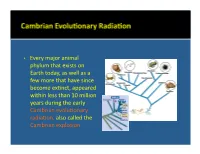
• Every Major Animal Phylum That Exists on Earth Today, As Well As A
• Every major animal phylum that exists on Earth today, as well as a few more that have since become ex:nct, appeared within less than 10 million years during the early Cambrian evolu:onary radiaon, also called the Cambrian explosion. • Phylum Pro:sta includes a diverse group of eukaryo:c microorganisms which are mostly unicellular. They are not necessarily primi:ve, although some are. Only some have a skeleton that can be preserved as a fossil. • Foraminifera and radiolaria are the most important examples. Coccolithophores, diatoms, and dinoflagellates are also examples. • Foraminifera are marine organisms that may be either planktonic, living in the water column where they float at various levels, or benthic, living on or within the seafloor sediment. • They form relavely large (0.1 mm to 5 cm) porous internal skeletons called tests through which project strands of living cytoplasm. The single- to mul:-chambered tests may be composed of organic maer, agglu:nated sand grains or sponge spicules, or calcium carbonate. • Benthic foraminifera • Planktonic foraminifera (Cambrian to Recent). (Jurassic to Recent). Oligocene Eocene • Radiolaria are planktonic marine organisms that tend to live in relavely cold water. • Their tests, oNen delicate and elaborate, are made of silica and presently accumulate on the floors of the parts of oceans that are deeper than those where foraminiferal tests are accumulang. • Radiolaria (Cambrian to • Radiolaria (Cambrian to Recent). Recent). Paleocene Radiolaria • Coccolithophores (Triassic to Recent) are extremely small marine planktonic photosynthe:c unicellular algae that are enclosed by plates of low-Mg calcite called coccoliths. Cretaceous White Cliffs of Dover, England • Diatoms (Jurassic to Recent) are small freshwater and marine planktonic photosynthe:c unicellular algae that are enclosed by a cell wall made of silica called a frustule. -

The Nervous System of the Lophophore in the Ctenostome Amathia Gracilis
Temereva and Kosevich BMC Evolutionary Biology (2016) 16:181 DOI 10.1186/s12862-016-0744-7 RESEARCH ARTICLE Open Access The nervous system of the lophophore in the ctenostome Amathia gracilis provides insight into the morphology of ancestral ectoprocts and the monophyly of the lophophorates Elena N. Temereva* and Igor A. Kosevich Abstract Background: The Bryozoa (=Ectoprocta) is a large group of bilaterians that exhibit great variability in the innervation of tentacles and in the organization of the cerebral ganglion. Investigations of bryozoans from different groups may contribute to the reconstruction of the bryozoan nervous system bauplan. A detailed investigation of the polypide nervous system of the ctenostome bryozoan Amathia gracilis is reported here. Results: The cerebral ganglion displays prominent zonality and has at least three zones: proximal, central, and distal. The proximal zone is the most developed and contains two large perikarya giving rise to the tentacle sheath nerves. The neuroepithelial organization of the cerebral ganglion is revealed. The tiny lumen of the cerebral ganglion is represented by narrow spaces between the apical projections of the perikarya of the central zone. The cerebral ganglion gives rise to five groups of main neurite bundles of the lophophore and the tentacle sheath: the circum-oral nerve ring, the lophophoral dorso-lateral nerves, the pharyngeal and visceral neurite bundles, the outer nerve ring, and the tentacle sheath nerves. Serotonin-like immunoreactive nerve system of polypide includes eight large perikarya located between tentacles bases. There are two analmost and six oralmost perikarya with prominent serotonergic “gap” between them. Based on the characteristics of their innervations, the tentacles can be subdivided into two groups: four that are near the anus and six that are near the mouth. -

Genes with Spiralian-Specific Protein Motifs Are Expressed In
ARTICLE https://doi.org/10.1038/s41467-020-17780-7 OPEN Genes with spiralian-specific protein motifs are expressed in spiralian ciliary bands Longjun Wu1,6, Laurel S. Hiebert 2,7, Marleen Klann3,8, Yale Passamaneck3,4, Benjamin R. Bastin5, Stephan Q. Schneider 5,9, Mark Q. Martindale 3,4, Elaine C. Seaver3, Svetlana A. Maslakova2 & ✉ J. David Lambert 1 Spiralia is a large, ancient and diverse clade of animals, with a conserved early developmental 1234567890():,; program but diverse larval and adult morphologies. One trait shared by many spiralians is the presence of ciliary bands used for locomotion and feeding. To learn more about spiralian- specific traits we have examined the expression of 20 genes with protein motifs that are strongly conserved within the Spiralia, but not detectable outside of it. Here, we show that two of these are specifically expressed in the main ciliary band of the mollusc Tritia (also known as Ilyanassa). Their expression patterns in representative species from five more spiralian phyla—the annelids, nemerteans, phoronids, brachiopods and rotifers—show that at least one of these, lophotrochin, has a conserved and specific role in particular ciliated structures, most consistently in ciliary bands. These results highlight the potential importance of lineage-specific genes or protein motifs for understanding traits shared across ancient lineages. 1 Department of Biology, University of Rochester, Rochester, NY 14627, USA. 2 Oregon Institute of Marine Biology, University of Oregon, Charleston, OR 97420, USA. 3 Whitney Laboratory for Marine Bioscience, University of Florida, 9505 Ocean Shore Blvd., St. Augustine, FL 32080, USA. 4 Kewalo Marine Laboratory, PBRC, University of Hawaii, 41 Ahui Street, Honolulu, HI 96813, USA. -

Phylum Porifera
790 Chapter 28 | Invertebrates updated as new information is collected about the organisms of each phylum. 28.1 | Phylum Porifera By the end of this section, you will be able to do the following: • Describe the organizational features of the simplest multicellular organisms • Explain the various body forms and bodily functions of sponges As we have seen, the vast majority of invertebrate animals do not possess a defined bony vertebral endoskeleton, or a bony cranium. However, one of the most ancestral groups of deuterostome invertebrates, the Echinodermata, do produce tiny skeletal “bones” called ossicles that make up a true endoskeleton, or internal skeleton, covered by an epidermis. We will start our investigation with the simplest of all the invertebrates—animals sometimes classified within the clade Parazoa (“beside the animals”). This clade currently includes only the phylum Placozoa (containing a single species, Trichoplax adhaerens), and the phylum Porifera, containing the more familiar sponges (Figure 28.2). The split between the Parazoa and the Eumetazoa (all animal clades above Parazoa) likely took place over a billion years ago. We should reiterate here that the Porifera do not possess “true” tissues that are embryologically homologous to those of all other derived animal groups such as the insects and mammals. This is because they do not create a true gastrula during embryogenesis, and as a result do not produce a true endoderm or ectoderm. But even though they are not considered to have true tissues, they do have specialized cells that perform specific functions like tissues (for example, the external “pinacoderm” of a sponge acts like our epidermis). -

Durham E-Theses
Durham E-Theses The palaeobiology of the panderodontacea and selected other euconodonts Sansom, Ivan James How to cite: Sansom, Ivan James (1992) The palaeobiology of the panderodontacea and selected other euconodonts, Durham theses, Durham University. Available at Durham E-Theses Online: http://etheses.dur.ac.uk/5743/ Use policy The full-text may be used and/or reproduced, and given to third parties in any format or medium, without prior permission or charge, for personal research or study, educational, or not-for-prot purposes provided that: • a full bibliographic reference is made to the original source • a link is made to the metadata record in Durham E-Theses • the full-text is not changed in any way The full-text must not be sold in any format or medium without the formal permission of the copyright holders. Please consult the full Durham E-Theses policy for further details. Academic Support Oce, Durham University, University Oce, Old Elvet, Durham DH1 3HP e-mail: [email protected] Tel: +44 0191 334 6107 http://etheses.dur.ac.uk The copyright of this thesis rests with the author. No quotation from it should be pubHshed without his prior written consent and information derived from it should be acknowledged. THE PALAEOBIOLOGY OF THE PANDERODONTACEA AND SELECTED OTHER EUCONODONTS Ivan James Sansom, B.Sc. (Graduate Society) A thesis presented for the degree of Doctor of Philosophy in the University of Durham Department of Geological Sciences, July 1992 University of Durham. 2 DEC 1992 Contents CONTENTS CONTENTS p. i ACKNOWLEDGMENTS p. viii DECLARATION AND COPYRIGHT p.