Кариотипическая Изменчивость И Систематика Серых Полевок (Rodentia, Arvicolini) Сообщение I
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mammal Species Native to the USA and Canada for Which the MIL Has an Image (296) 31 July 2021
Mammal species native to the USA and Canada for which the MIL has an image (296) 31 July 2021 ARTIODACTYLA (includes CETACEA) (38) ANTILOCAPRIDAE - pronghorns Antilocapra americana - Pronghorn BALAENIDAE - bowheads and right whales 1. Balaena mysticetus – Bowhead Whale BALAENOPTERIDAE -rorqual whales 1. Balaenoptera acutorostrata – Common Minke Whale 2. Balaenoptera borealis - Sei Whale 3. Balaenoptera brydei - Bryde’s Whale 4. Balaenoptera musculus - Blue Whale 5. Balaenoptera physalus - Fin Whale 6. Eschrichtius robustus - Gray Whale 7. Megaptera novaeangliae - Humpback Whale BOVIDAE - cattle, sheep, goats, and antelopes 1. Bos bison - American Bison 2. Oreamnos americanus - Mountain Goat 3. Ovibos moschatus - Muskox 4. Ovis canadensis - Bighorn Sheep 5. Ovis dalli - Thinhorn Sheep CERVIDAE - deer 1. Alces alces - Moose 2. Cervus canadensis - Wapiti (Elk) 3. Odocoileus hemionus - Mule Deer 4. Odocoileus virginianus - White-tailed Deer 5. Rangifer tarandus -Caribou DELPHINIDAE - ocean dolphins 1. Delphinus delphis - Common Dolphin 2. Globicephala macrorhynchus - Short-finned Pilot Whale 3. Grampus griseus - Risso's Dolphin 4. Lagenorhynchus albirostris - White-beaked Dolphin 5. Lissodelphis borealis - Northern Right-whale Dolphin 6. Orcinus orca - Killer Whale 7. Peponocephala electra - Melon-headed Whale 8. Pseudorca crassidens - False Killer Whale 9. Sagmatias obliquidens - Pacific White-sided Dolphin 10. Stenella coeruleoalba - Striped Dolphin 11. Stenella frontalis – Atlantic Spotted Dolphin 12. Steno bredanensis - Rough-toothed Dolphin 13. Tursiops truncatus - Common Bottlenose Dolphin MONODONTIDAE - narwhals, belugas 1. Delphinapterus leucas - Beluga 2. Monodon monoceros - Narwhal PHOCOENIDAE - porpoises 1. Phocoena phocoena - Harbor Porpoise 2. Phocoenoides dalli - Dall’s Porpoise PHYSETERIDAE - sperm whales Physeter macrocephalus – Sperm Whale TAYASSUIDAE - peccaries Dicotyles tajacu - Collared Peccary CARNIVORA (48) CANIDAE - dogs 1. Canis latrans - Coyote 2. -

Status of Birds of Oak Creek Wildlife Area
Status of Birds of Oak Creek Wildlife Area Abundance Seasonal Occurance *Species range included in C = Common r= Resident Oak Creek WLA but no U = Uncommon s = Summer Visitor documentation of species R = Rare w= Winter Visitor m=Migrant Common Name Genus Species Status Common Loon* Gavia immer Rw Pied-billed Grebe Podilymbus podiceps Cr Horned Grebe Podiceps auritus Um Eared Grebe Podiceps nigricollis Us Western Grebe Aechmorphorus occidentalis Us Clark's Grebe* Aechmorphorus clarkii Rm Double-crested Cormorant Phalocrorax auritus Um American Bittern* Botaurus lentiginosus Us Great Blue Heron Arden herodias Cr Black-crowned Night-Heron Nycticorax nycticorax Cr Tundra Swan Cygnus columbiaus Rw Trumpeter Swan Cygnus buccinator Am Greater White-fronted Anser albifrons Rm Goose* Snow Goose Chen caerulescens Rw Canada Goose Branta canadensis Cr Green-winged Teal Anas crecca Ur Mallard Anas Platyrynchos Cr Northen Pintail Anas acuta Us Blue-winged Teal Anas discors Rm Cinnamon Teal Anas cyanoptera Us Northern Shoveler Anas clypeata Cr Gadwall* Anas strepere Us Eurasian Wigeon* Anas Penelope Rw American Wigeon Anas Americana Cr Wood Duck Aix sponsa Ur Redhead Aythya americana Uw Canvasback Aythya valisineria Uw Ring-necked Duck Aythya collaris Uw Greater Scaup* Aythya marila Rw Leser Scaup Aythya affinis Uw Common Goldeneye Bucephala clangula Uw Barrow's Goldeneye Bucephala islandica Rw Bufflehead Bucephala albeola Cw Harlequin Duck Histrionicus histrionicus Rs White-winged Scoter Melanitta fusca Rm Hooded Merganser Lophodytes Cucullatus Rw -

MAMMALS of WASHINGTON Order DIDELPHIMORPHIA
MAMMALS OF WASHINGTON If there is no mention of regions, the species occurs throughout the state. Order DIDELPHIMORPHIA (New World opossums) DIDELPHIDAE (New World opossums) Didelphis virginiana, Virginia Opossum. Wooded habitats. Widespread in W lowlands, very local E; introduced from E U.S. Order INSECTIVORA (insectivores) SORICIDAE (shrews) Sorex cinereus, Masked Shrew. Moist forested habitats. Olympic Peninsula, Cascades, and NE corner. Sorex preblei, Preble's Shrew. Conifer forest. Blue Mountains in Garfield Co.; rare. Sorex vagrans, Vagrant Shrew. Marshes, meadows, and moist forest. Sorex monticolus, Montane Shrew. Forests. Cascades to coast, NE corner, and Blue Mountains. Sorex palustris, Water Shrew. Mountain streams and pools. Olympics, Cascades, NE corner, and Blue Mountains. Sorex bendirii, Pacific Water Shrew. Marshes and stream banks. W of Cascades. Sorex trowbridgii, Trowbridge's Shrew. Forests. Cascades to coast. Sorex merriami, Merriam's Shrew. Shrub steppe and grasslands. Columbia basin and foothills of Blue Mountains. Sorex hoyi, Pygmy Shrew. Many habitats. NE corner (known only from S Stevens Co.), rare. TALPIDAE (moles) Neurotrichus gibbsii, Shrew-mole. Moist forests. Cascades to coast. Scapanus townsendii, Townsend's Mole. Meadows. W lowlands. Scapanus orarius, Coast Mole. Most habitats. W lowlands, central E Cascades slopes, and Blue Mountains foothills. Order CHIROPTERA (bats) VESPERTILIONIDAE (vespertilionid bats) Myotis lucifugus, Little Brown Myotis. Roosts in buildings and caves. Myotis yumanensis, Yuma Myotis. All habitats near water, roosting in trees, buildings, and caves. Myotis keenii, Keen's Myotis. Forests, roosting in tree cavities and cliff crevices. Olympic Peninsula. Myotis evotis, Long-eared Myotis. Conifer forests, roosting in tree cavities, caves and buildings; also watercourses in arid regions. -

Life History Account for Creeping Vole
California Wildlife Habitat Relationships System California Department of Fish and Wildlife California Interagency Wildlife Task Group CREEPING VOLE Microtus oregoni Family: MURIDAE Order: RODENTIA Class: MAMMALIA M137 Written by: P. Brylski, J. Harris Reviewed by: H. Shellhammer Edited by: R. Duke, S. Granholm DISTRIBUTION, ABUNDANCE, AND SEASONALITY In California, the creeping vole is found in the northwest corner of the state from Del Norte and Siskiyou cos. south to Mendocino Co., including the extreme western portions of Tehama and Shasta cos. Common to abundant in herbaceous and shrubby understories of riparian and coniferous forests, including burned and clearcut areas. Also common at times in grassland and wet meadow habitats (Goertz 1964, Gashwiler 1972), as well as cropland. SPECIFIC HABITAT REQUIREMENTS Feeding: Feeds on grasses and forbs from ground surface, and some underground fungi (Maser et al. 1978). Cover: Seeks cover in ground litter, dense herbaceous undergrowth, and in burrows and log cavities. Burrows are excavated in soft soil. Occasionally, the burrows of other small mammals (particularly moles) are used. Reproduction: A nest of dried grass is constructed in burrow. Water: Probably does not require free water. Pattern: Prefers herbaceous habitat or shrublands with herbaceous openings, with soft soil for burrowing. Early successional stages of burned and cutover areas at the edge of coniferous forests support the highest densites. SPECIES LIFE HISTORY Activity Patterns: Yearlong activity. Mainly nocturnal, but probably forages near nest throughout the day. Seasonal Movements/Migration: None. Home Range: In Oregon clearcuts, home ranges averaged 0.06 ha (0.14 ac) for males, and 0.09 ha (0.22 ac) for females (Gashwiler 1972). -

Regulation of Sex Chromosome Constitution of Somatic and Germ Cells in the Wood Lemming A
Regulation of sex chromosome constitution of somatic and germ cells in the wood lemming A. Gropp, K. Fredga, H. Winking, F. Frank To cite this version: A. Gropp, K. Fredga, H. Winking, F. Frank. Regulation of sex chromosome constitution of somatic and germ cells in the wood lemming. Annales de biologie animale, biochimie, biophysique, 1978, 18 (2B), pp.367-375. hal-00897299 HAL Id: hal-00897299 https://hal.archives-ouvertes.fr/hal-00897299 Submitted on 1 Jan 1978 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Regulation of sex chromosome constitution of somatic and germ cells in the wood lemming A. GROPP K. FREDGA H. WINKING, F. FRANK Abteilung für Pathologie, Medizinische Hochschule Lü6eck Ratzeburger Allee 160, Fed. Rep. Germany. * Institute of Genetics, University of Lund, S-223 62 Lund, Sweden. ** Biologische Bundesanstalt, Messeweg 11/12, 3300 Braunschweig, Fed. Rep. Germany. Summary. The wood lemming displays certain peculiar features. (a) The sex ratio shows a prevalence of females, and some females produce only female offspring. (b) A conside- rable proportion of the females has XY sex chromosomes in the somatic tissues, but the Y is absent in the germ line. -

Complete List of Amphibian, Reptile, Bird and Mammal Species in California
Complete List of Amphibian, Reptile, Bird and Mammal Species in California California Department of Fish and Game Sept. 2008 (updated) This list represents all of the native or introduced amphibian, reptile, bird and mammal species known in California. Introduced species are marked with “I”, harvest species with “HA”, and vagrant species or species with extremely limited distributions with *. The term “introduced”, as used here, represents both accidental and intentional introductions. Subspecies are not included on this list. The most current list of species and subspecies with special management status is available from the California Natural Diversity Database (CNDDB) Taxonomy and nomenclature used within the list are the same as those used within both the CNDDB and CWHR software programs and data sets. If a discrepancy exists between this list and the ones produced by CNDDB, the CNDDB list can be presumed to be more accurate as it is updated more frequently than the CWHR data set. ________________________________________________________________________ ______________________________________________________________________ ______________________________________________________________________ AMPHIBIA (Amphibians) CAUDATA (Salamanders) AMBYSTOMATIDAE (Mole Salamanders and Relatives) Long-toed Salamander Ambystoma macrodactylum Tiger Salamander Ambystoma tigrinum I California Tiger Salamander Ambystoma californiense Northwestern Salamander Ambystoma gracile RHYACOTRITONIDAE (Torrent or Seep Salamanders) Southern Torrent Salamander Rhyacotriton -

Information to Users
Molecular phylogenetics of arvicoline rodents Item Type Thesis Authors Conroy, Christopher John Download date 24/09/2021 20:10:57 Link to Item http://hdl.handle.net/11122/9519 INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type o f computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. Each original is also photographed in one exposure and is included in reduced form at the back o f the book. Photographs included in the original manuscript have been reproduced xerographically in this copy. Higher quality 6” x 9” black and white photographic prints are available for any photographs or illustrations appearing in this copy for an additional charge. Contact UMI directly to order. UMI A Bell & Howell Information Company 300 North Zeeb Road, Ann Arbor MI 48106-1346 USA 313/761-4700 800/521-0600 Reproduced with permission of the copyright owner. -

List of Taxa for Which MIL Has Images
LIST OF 27 ORDERS, 163 FAMILIES, 887 GENERA, AND 2064 SPECIES IN MAMMAL IMAGES LIBRARY 31 JULY 2021 AFROSORICIDA (9 genera, 12 species) CHRYSOCHLORIDAE - golden moles 1. Amblysomus hottentotus - Hottentot Golden Mole 2. Chrysospalax villosus - Rough-haired Golden Mole 3. Eremitalpa granti - Grant’s Golden Mole TENRECIDAE - tenrecs 1. Echinops telfairi - Lesser Hedgehog Tenrec 2. Hemicentetes semispinosus - Lowland Streaked Tenrec 3. Microgale cf. longicaudata - Lesser Long-tailed Shrew Tenrec 4. Microgale cowani - Cowan’s Shrew Tenrec 5. Microgale mergulus - Web-footed Tenrec 6. Nesogale cf. talazaci - Talazac’s Shrew Tenrec 7. Nesogale dobsoni - Dobson’s Shrew Tenrec 8. Setifer setosus - Greater Hedgehog Tenrec 9. Tenrec ecaudatus - Tailless Tenrec ARTIODACTYLA (127 genera, 308 species) ANTILOCAPRIDAE - pronghorns Antilocapra americana - Pronghorn BALAENIDAE - bowheads and right whales 1. Balaena mysticetus – Bowhead Whale 2. Eubalaena australis - Southern Right Whale 3. Eubalaena glacialis – North Atlantic Right Whale 4. Eubalaena japonica - North Pacific Right Whale BALAENOPTERIDAE -rorqual whales 1. Balaenoptera acutorostrata – Common Minke Whale 2. Balaenoptera borealis - Sei Whale 3. Balaenoptera brydei – Bryde’s Whale 4. Balaenoptera musculus - Blue Whale 5. Balaenoptera physalus - Fin Whale 6. Balaenoptera ricei - Rice’s Whale 7. Eschrichtius robustus - Gray Whale 8. Megaptera novaeangliae - Humpback Whale BOVIDAE (54 genera) - cattle, sheep, goats, and antelopes 1. Addax nasomaculatus - Addax 2. Aepyceros melampus - Common Impala 3. Aepyceros petersi - Black-faced Impala 4. Alcelaphus caama - Red Hartebeest 5. Alcelaphus cokii - Kongoni (Coke’s Hartebeest) 6. Alcelaphus lelwel - Lelwel Hartebeest 7. Alcelaphus swaynei - Swayne’s Hartebeest 8. Ammelaphus australis - Southern Lesser Kudu 9. Ammelaphus imberbis - Northern Lesser Kudu 10. Ammodorcas clarkei - Dibatag 11. Ammotragus lervia - Aoudad (Barbary Sheep) 12. -

Washington Species List
TheThe NatureMappingNatureMapping ProgramProgram Washington Cooperative Fish and Wildlife Research Unit School of Fisheries and Aquatic Sciences University of Washington Species of Washington • Amphibians • Reptiles • Fish • Birds • Mammals For more information on NatureMapping http://depts.washington.edu/natmap last update: 01/03/06 The NatureMapping Program Amphibians (27 species) Frogs/Toads (13 species) RA99 Unidentified Rana Rana sp. RACAT Bullfrog Rana catesbeiana RANCA Cascades frog Rana cascadae SPIN Great basin spadefoot Scaphiopus intermontana RACL Green frog Rana clamitans RAPI Northern leopard frog Rana pipiens PSRE Pacific treefrog (Chorus frog) Hyla regilla RAAU Red-legged frog Rana aurora RALU Spotted frog (Eastside) Rana luteiventris RAPR Spotted frog (Westside) Rana pretiosa ASTR Tailed frog Ascaphus truei BUBO Western toad Bufo boreas RASY Wood frog Rana sylvatica BUWO Woodhouse's toad Bufo woodhousei Salamanders/Newts (14 species) AM99 Unidentified Ambystoma Ambystoma sp. RHCAS Cascade torrent salamander Rhyacotriton cascadae RHKE Columbia torrent salamander Rhyacotriton kezeri DICO Cope's giant salamander Dicamptodon copei PLDU Dunn's salamander Plethodon dunni ENES Ensatina Ensatina eschscholtzi PLLA Larch mountain salamander Plethodon larselli AMMA Long-toed salamander Ambystoma macrodactylum AMGR Northwestern salamander Ambystoma gracile RHOL Olympic torrent salamander Rhyacotriton olympicus DITE Pacific giant salamander Dicamptodon tenebrosus TAGR Roughskin newt Taricha granulosa AMTI Tiger salamander Ambystoma -

Appendix C Wildlife Supplement
Appendix C Wildlife Supplement Contents Table 1. Land protection status by habitat type Table 2. Species occurrences for the Hood Subbasin based on current wildlife habitat types (IBIS) Table 1. Land Protection Status by habitat type using GIS data layers provided by the Northwest Habitat Institute for the subbasin planning effort. Total Acres subject to discrepancies in input data layers. Metadata link is at www.nwhi.org/ibis/mapping/gisdata/docs/crb/crb_lsstatus_meta.htm. Analysis Area Habitat Protection Status Acres Columbia River Tributaries 63022 Agriculture, pasture and mixed environs 2382 Low 6 Medium 4 None 2372 Montane mixed conifer forest 3653 High 813 Low 2660 Medium 89 None 91 Open water - lakes, rivers, streams 152 High 48 Low 43 Medium 26 None 35 Ponderosa pine forest and woodlands 2 High 2 Urban and mixed environs 1280 Low 126 None 1154 Westside lowlands conifer-hardwood forest 55553 High 31236 Low 17977 Medium 1226 None 5114 Hood River Basin 217492 Agriculture, pasture and mixed environs 33400 Low 152 None 33248 Alpine grassland and shrublands 4469 High 3386 Low 1083 Eastside (interior) grasslands 1538 Low 169 None 1369 Eastside (interior) mixed conifer forest 23194 Low 8252 None 14942 Herbaceous wetlands 197 High 56 Low 141 Montane coniferous wetlands 116 Low 99 None 16 Montane mixed conifer forest 47895 High 14560 Low 33061 None 274 Open water - lakes, rivers, streams 523 High 250 Low 217 None 55 Ponderosa pine forest and woodlands 4739 High 13 Low 1341 None 3385 Subalpine parkland 4394 High 2883 Low 1511 Urban and mixed environs 763 None 763 Westside lowlands conifer-hardwood forest 95388 High 2357 Low 51835 None 41196 Westside oak and dry Douglas-fir forest and woo 876 Low 240 None 636 Subbasin Species Occurrences Generated by IBIS on 10/21/2003 11:09:13 AM. -

Oregon Department of Fish and Wildlife
Oregon Department of Fish and Wildlife Wildlife Control Operator Training Manual Revised March 31, 2016 TABLE OF CONTENTS Part 1 Page # Introduction 5 Wildlife “Damage” and Wildlife “Nuisance” 5 Part 2 History 8 Definitions 9 Rules and Regulations 11 Part 3 Business Practices 13 Part 4 Methods of Control 15 Relocation of Wildlife 16 Handling of Sick or Injured Wildlife 17 Wildlife That Has Injured a Person 17 Euthanasia 18 Transportation 19 Disposal of Wildlife 20 Migratory Birds 20 Diseases (transmission, symptoms, human health) 20 Part 5 Liabilities 26 Record Keeping and Reporting Requirements 26 Live Animals and Sale of Animals 26 Complaints 27 Cancellation or Non-Renewal of Permit 27 Part 6 Best Management Practices 29 Traps 30 Bodygrip Traps 30 Foothold Traps 30 Box Traps 31 Snares 31 Specialty Traps 32 Trap Sets 33 Bait and Lures 33 Equipment 33 Trap Maintenance and Safety 34 Releasing Nontarget Wildlife 34 2 Resources 36 Part 7 Species: Bats 38 Beaver 47 Coyote 49 Eastern Cottontail 52 Eastern Gray Squirrel 54 Eastern Fox Squirrel 56 Western (Silver) Gray Squirrel 58 Gray Fox 60 Red Fox 62 Mountain Beaver 64 Muskrat 66 Nutria 68 Opossum 70 Porcupine 72 Raccoon 74 Striped Skunk 77 Oregon Species List 79 Part 8 Acknowledgements 83 References and Resources 83 3 Part 1 Introduction Wildlife “Damage” and Wildlife “ Nuisance” 4 Introduction As Oregon becomes more urbanized, wildlife control operators are playing an increasingly important role in wildlife management. The continued development and urbanization of forest and farmlands is increasing the chance for conflicts between humans and wildlife. -
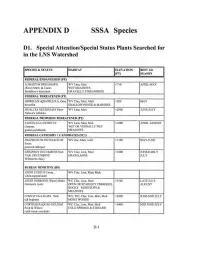
APPENDIXD SSSA Species
APPENDIXD SSSA Species Dl. Special Attention/Special Status Plants Searched for in the LNS Watershed SPECIES & STATUS HABITAT ELEVATION BEST I.D. (Ff) SEASON FEDERAL ENDANGERED (FE) LOMATJUM BRADSHAW!! WV Linn, Mari <750 APRIL-MAY (Rose) Math. & Const. WET MEADOWS Bradshaw's lomatium GRAVELLY STREAMBEDS FEDERALTHREATENED(FT) HOWELL/A AQUATJLL/S A. Grdy WV Clac, Mari, Mult <200 MAY howe Ilia SHALLOW PONDS & MARSHES SJDALCEA NELSON/ANA Piper WV Linn, Mari <2000 JUNE-JULY Nelson's sidalcea FEDERAL PROPOSED THREATENED (PT) CASTILLEJA I~EVJSECTA WV Linn, Mari, Mull <1000 APRJL..A UGUST Greenm. WET OR VERNALLY WET golden paintbrush MEADOWS FEDERAL CATEGORY 1 CANDIDATES (FCl) Dl:.'LPHLNJUM PAVONACEUM WV clac, Mari, mult < 1500 MAY-JUNE Ewan peacock larkspur ERIGERON fJECVMBENS Nutt. WV Clac, Linn, Mari < 1000 JUNE-EARLY VAR . DECVMBENS GRASSLANDS JULY Willamette daisy BUREAU SENSITIVE (BS) ASTER CURTUS Cronq. WV Clac. Linn, Mari, Mult white-topped aster ASTER GORMAN!! (Piper) Blake WC Clac, Linn, Mari >3500 LATE JULY- Gorman's Aster OPEN OR SPARSLEY TIMBERED, AUGUST ROCKY RIDGETOPS & MEADOWS CIMTOFUGA ELATA Noll. WV, WC, Clac, Linn, Mari, Mull <2000 JUNE-MID JULY tall bugbane MOIST WOODS CORYDALIS AQUAE-GEUDAE WC Clac, Linn, Mari, Mult > 1000 M[O JUNE-JULY Peck&Wilson COLD SPRINGS & STREAMS cold-water corydalis D-1 IS PECIES & STATUS II HABITAT II ~VATION IIB EST I.D . .. SEASON I DELP!flNJUM LEUCAPFIAEUM WV Clac, Mari, Mull <1000 MAY-EARLY Greene JUNE white rock larkspur DELPHINIUM OREGANUM WV Linn, Marl LOW 1/ow. Willamette Valley larkspur HOR.KELIA CONGESTA Douglas WVLinn ww APRIL-JUNE ssp. CONGESTA OPEN SANDY OR ROCKY FLATS shaggy borkelia TO OPEN WOODS LVP1NUS SULPHUREUS WV Linn, Mari <1500 MAY-JULY Douglas ssp.