Defects in Vestibular Sensory Epithelia and Innervation in Mice with Loss of Chd7 Function: Implications for Human CHARGE Syndrome
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
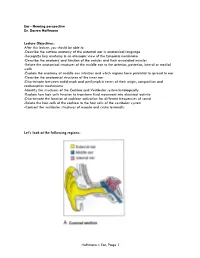
Ear, Page 1 Lecture Outline
Ear - Hearing perspective Dr. Darren Hoffmann Lecture Objectives: After this lecture, you should be able to: -Describe the surface anatomy of the external ear in anatomical language -Recognize key anatomy in an otoscopic view of the tympanic membrane -Describe the anatomy and function of the ossicles and their associated muscles -Relate the anatomical structures of the middle ear to the anterior, posterior, lateral or medial walls -Explain the anatomy of middle ear infection and which regions have potential to spread to ear -Describe the anatomical structures of the inner ear -Discriminate between endolymph and perilymph in terms of their origin, composition and reabsorption mechanisms -Identify the structures of the Cochlea and Vestibular system histologically -Explain how hair cells function to transform fluid movement into electrical activity -Discriminate the location of cochlear activation for different frequencies of sound -Relate the hair cells of the cochlea to the hair cells of the vestibular system -Contrast the vestibular structures of macula and crista terminalis Let’s look at the following regions: Hoffmann – Ear, Page 1 Lecture Outline: C1. External Ear Function: Amplification of Sound waves Parts Auricle Visible part of external ear (pinna) Helix – large outer rim Tragus – tab anterior to external auditory meatus External auditory meatus Auditory Canal/External Auditory Meatus Leads from Auricle to Tympanic membrane Starts cartilaginous, becomes bony as it enters petrous part of temporal bone Earwax (Cerumen) Complex mixture -

Mathematical Model of the Cupula-Endolymph System with Morphological Parameters for the Axolotl (Ambystoma Tigrinum) Semicircular Canals
138 The Open Medical Informatics Journal, 2008, 2, 138-148 Open Access Mathematical Model of the Cupula-Endolymph System with Morphological Parameters for the Axolotl (Ambystoma tigrinum) Semicircular Canals Rosario Vega1, Vladimir V. Alexandrov2,3, Tamara B. Alexandrova1,3 and Enrique Soto*,1 1Instituto de Fisiología, Universidad Autónoma de Puebla, 2Facultad de Ciencias Físico Matemáticas, Universidad Autónoma de Puebla, 3 Lomonosov Moscow State University, Mexico Abstract: By combining mathematical methods with the morphological analysis of the semicircular canals of the axolotl (Ambystoma tigrinum), a system of differential equations describing the mechanical coupling in the semicircular canals was obtained. The coefficients of this system have an explicit physiological meaning that allows for the introduction of morphological and dynamical parameters directly into the differential equations. The cupula of the semicircular canals was modeled both as a piston and as a membrane (diaphragm like), and the duct canals as toroids with two main regions: i) the semicircular canal duct and, ii) a larger diameter region corresponding to the ampulla and the utricle. The endolymph motion was described by the Navier-Stokes equations. The analysis of the model demonstrated that cupular behavior dynamics under periodic stimulation is equivalent in both the piston and the membrane cupular models, thus a general model in which the detailed cupular structure is not relevant was derived. Keywords: Inner ear, vestibular, hair cell, transduction, sensory coding, physiology. 1. INTRODUCTION linear acceleration detectors, and the SCs as angular accel- eration detectors, notwithstanding that both sensory organs The processing of sensory information in the semicircular are based on a very similar sensory cell type. -

Organum Vestibulocochleare INTERNAL EAR MIDDLE EAR EXTERNAL EAR PETROSAL BONE- Eq EXTERNAL EAR AURICLE
EAR organum vestibulocochleare INTERNAL EAR MIDDLE EAR EXTERNAL EAR PETROSAL BONE- Eq EXTERNAL EAR AURICLE The external ear plays the role of an acoustic antenna: auricle the auricle (together with the head) collects and focuses sound waves, the ear canal act as a resonator. tympanic membrane anular cartilage meatus acusticus externus EXTERNAL EAR EXTERNAL EAR AURICLE scutiform cartilage Auricular muscles: -Dorsal -Ventral -Rostral -Caudal EXTERNAL EAR MEATUS ACUSTICUS EXTERNUS auricular cartilage vertical canal auditory ossicles horizontal cochlea canal auditory tube tympanic tympanic eardrum bulla cavity tympanic membrane MIDDLE EAR Auditory ossicles STAPES INCUS Tympanic cavity: (anvil) (stirrup) - epitympanium - mesotympanium - hypotympanium MALLEUS (hammer) auditory vestibular window- ossicles or oval window through which mechanical stimuli (transmitted by the auditory ossicles) enter the epitympanic internal ear for translation recess into nerve impulses auditory tube (Eustachian tube) cochlear window- or round window tympanic cavity bulla tympanica through which the vibration of the perilympha is absorbed MIDDLE EAR MIDDLE EAR GUTTURAL POUCH- Eq MIDDLE EAR AUDITORY OSSICLES head INCUS processus rostralis (stirrup) STAPES processus muscularis (anvil) manubrium short crus body MALLEUS (hammer) Two muscles of the ossicles: long crus m. tensor tympani- n. tensoris tympani ex. n. base mandibularis (footplate) m. stapedius- n. stapedius ex. n. facialis crus The muscles fix the bones and protect the cochlea crus against the harmful effects -

A Place Principle for Vertigo
Auris Nasus Larynx 35 (2008) 1–10 www.elsevier.com/locate/anl A place principle for vertigo Richard R. Gacek * Department of Otolaryngology, Head and Neck Surgery, University of Massachusetts Medical School, Worcester, MA 01655, USA Received 16 March 2007; accepted 13 April 2007 Available online 24 October 2007 Abstract Objective: To provide a road map of the vestibular labyrinth and its innervation leading to a place principle for different forms of vertigo. Method: The literature describing the anatomy and physiology of the vestibular system was reviewed. Results: Different forms of vertigo may be determined by the type of sense organ, type of ganglion cell and location in the vestibular nerve. Conclusion: Partial lesions (viral) of the vestibular ganglion are manifested as various forms of vertigo. # 2007 Elsevier Ireland Ltd. All rights reserved. Keywords: Vertigo; Vestibular nerve; Pathology Contents 1. Introduction . ............................................................................... 1 2. Sense organ. ............................................................................... 2 3. Ganglion cells ............................................................................... 4 4. Hair cells . ............................................................................... 5 5. Hair cell polarization . ....................................................................... 5 6. Efferent vestibular system ....................................................................... 8 7. A place principle for vertigo . ................................................................. -

Mechanoreception
Mechanoreception Introduction Hair cells : the basic mechanosensory unit Hair cell structure Inner ear and accessory organ structures Vestibule Otolith organs Weberian ossicles Lateral line Lateral line structure Receptor organs Acoustic communication: sound production and reception Sound production mechanisms Locomotion and posture Introduction A mechanoreceptor is a sensory receptor that responds to mechanical pressure or distortion. In fishes mechanoreception concerns the inner ear and the lateral line system. Hair cells are the UNIVERSAL MECHANOSENSORY TRANSDUCERS in both the lateral line and hearing systems. The INNER EAR is responsible for fish EQUILIBRIUM, BALANCE and HEARING LATERAL LINE SYSTEM detects DISTURBANCES in the water. Hair cell structure EACH HAIR CELL CONSISTS OF TWO TYPES OF "HAIRS" OR RECEPTOR PROCESSES: Many microvillar processes called STEREOCILIA. One true cilium called the KINOCILLIUM. COLLECTIVELY, the cluster is called a CILIARY BUNDLE. The NUMBER OF STEREOCILIA PER BUNDLE IS VARIABLE, and ranges from a 10s of stereocilia to more than a 100. The STEREOCILIA PROJECT into a GELATINOUS CUPULA ON THE APICAL (exposed) SURFACE of the cell. The cilium and villi are ARRANGED IN A STEPWISE GRADATION - the longest hair is the kinocillium, and next to it, the stereocilia are arranged in order of decreasing length. These cells SYNAPSE WITH GANGLION CELLS. They have DIRECTIONAL PROPERTIES - response to a stimulus depends on the direction in which the hairs are bent. So, if the displacement causes the stereocilia to bend towards the kinocilium, the cell becomes DEPOLARIZED = EXCITATION. If the stereocilia bend in the opposite direction, the cell becomes HYPERPOLARIZED = INHIBITION of the cell. If the hair bundles are bent at a 90o angle to the axis of the kinocilium and stereocilia there will be no response. -

Novel Cell Types and Developmental Lineages Revealed by Single-Cell
RESEARCH ARTICLE Novel cell types and developmental lineages revealed by single-cell RNA-seq analysis of the mouse crista ampullaris Brent A Wilkerson1,2†, Heather L Zebroski1,2, Connor R Finkbeiner1,2, Alex D Chitsazan1,2,3‡, Kylie E Beach1,2, Nilasha Sen1, Renee C Zhang1, Olivia Bermingham-McDonogh1,2* 1Department of Biological Structure, University of Washington, Seattle, United States; 2Institute for Stem Cells and Regenerative Medicine, University of Washington, Seattle, United States; 3Department of Biochemistry, University of Washington, Seattle, United States Abstract This study provides transcriptomic characterization of the cells of the crista ampullaris, sensory structures at the base of the semicircular canals that are critical for vestibular function. We performed single-cell RNA-seq on ampullae microdissected from E16, E18, P3, and P7 mice. Cluster analysis identified the hair cells, support cells and glia of the crista as well as dark cells and other nonsensory epithelial cells of the ampulla, mesenchymal cells, vascular cells, macrophages, and melanocytes. Cluster-specific expression of genes predicted their spatially restricted domains of *For correspondence: gene expression in the crista and ampulla. Analysis of cellular proportions across developmental [email protected] time showed dynamics in cellular composition. The new cell types revealed by single-cell RNA-seq Present address: †Department could be important for understanding crista function and the markers identified in this study will of Otolaryngology-Head and enable the examination of their dynamics during development and disease. Neck Surgery, Medical University of South Carolina, Charleston, United States; ‡CEDAR, OHSU Knight Cancer Institute, School Introduction of Medicine, Portland, United States The vertebrate inner ear contains mechanosensory organs that sense sound and balance. -

NTID HONORS ANATOMY & PHYSIOLOTY – Mr. Barnett Honors
NTID HONORS ANATOMY & PHYSIOLOTY – Mr. Barnett Honors Anatomy & Physiology: Wednesday – Friday Read pp. 463 – 466 concerning Equilibrium and the corresponding notes attached Complete the Coloring Sheet over the Inner Ear attached (due Test day) **There is no Multiple Choice set of questions as was first indicated Book assignment and Multiple Choice review is due Monday HONORS ANATOMY & PHYSIOLOGY I. Equilibrium A. Equilibrium is primarily centered in the inner ear (vestibule and semicircular canals) B. Equilibrium is also regulated by vision 1. We use information that we see to determine much of our equilibrium—such as the sensation of “down” and “up” 2. Sometimes, conflicting information from our inner ear and eyes can result in our body thinking we’ve been poisoned, which activates the nausea centers of the medulla oblongata: motion sicKness a. Imagine riding in a car on a straight, smooth highway and you are looKing out the side window at the rushing trees passing by: i. Your inner ear detects no major changes in your inertial movement and thus thinKs you are motionless ii. Your eyes, however, see that you are moving iii. This discrepancy between the senses results in motion sicKness b. Imagine riding in a car on a winding, bumpy road as you try to read a book: i. Your inner ear detects your movements as you sway bacK and forth ii. Your eyes focused upon a single page, gives the sensation that you are motionless iii. This discrepancy between the senses results in motion sicKness C. Two types of Equilibrium: static and dynamic D. Static equilibrium 1. -

Proprioceptors
Proprioceptors equilibrium and balance are maintained by complex interactions between simple receptors and complex proprioceptor organs and vision simple receptors embedded in deep tissues; muscles, tendons, joints, etc Pacinian corpuscles muscle spindle fibers Golgi Tendon organs joint receptors they monitor slight changes in pressure as we shift positions stretching of various internal organs positions of limbs wrt rest of body more complex proprioceptor organs are found in the inner ear as part of special senses inner ear is a multiple sense organ in addition to hearing also has receptors for equilibrium static equilibrium orientation wrt gravity: position, orientation dynamic equilibrium changes in movement located in “vestibular apparatus” of inner ear otolith organs utricle – linear acceleration ?ck? saccule – rotational acceleration ?ck? semicircular canals part of membranous labyrinth Static Equilibrium within vestibule of inner ear is fluid filled sac = utricle contains patches of hair cells = macula embedded in gelatinous matrix (membrane) Human Anatomy & Physiology: Senses; Ziser, 2004 1 also in gelatinous material are small particles of calcium carbonate = otoliths as head moves these otoliths shift positions and stimulate different hair cells tells position and orientation Dynamic Equilibrium semicircular canals branch off vestibule fluid filled each canal is oriented along a different plane at right angles to each other at bases of each are swelling = ampulla each ampulla contains a patch of hair cells = crista ampullaris movement of fluid in these canals causes bending of crista ampullaris detect acceleration and deceleration detect turning, changes in motion detect direction of the change continuous movement in same direction cannot be detected eg. we are spinning on surface of earth at ~1000 mph eg. -

The Special Senses the Ear External Ear Middle
1/24/2016 The Ear • The organ of hearing and equilibrium – Cranial nerve VIII - Vestibulocochlear – Regions The Special Senses • External ear • Middle ear Hearing and • Internal ear (labyrinth) Equilibrium External Ear Middle Internal ear • Two parts External ear (labyrinth) ear – Pinna or auricle (external structures) – External auditory meatus (car canal) Auricle • Site of cerumen (earwax) production (pinna) – Waterproofing, protection • Separated from the middle ear by the tympanic membrane Helix (eardrum) – Vibrates in response to sound waves Lobule External acoustic Tympanic Pharyngotympanic meatus membrane (auditory) tube (a) The three regions of the ear Figure 15.25a Middle Ear Epitympanic Middle Ear Superior Malleus Incus recess Lateral • Tympanic cavity Anterior – Air-filled chamber – Openings View • Tympanic membrane – covers opening to outer ear • Round and oval windows – openings to inner ear • Epitympanic recess – dead-end cavity into temporal bone of unknown function • Auditory tube – AKA Eustachian tube or pharyngotympanic tube Pharyngotym- panic tube Tensor Tympanic Stapes Stapedius tympani membrane muscle muscle (medial view) Figure 15.26 1 1/24/2016 Middle Ear Middle Ear • Auditory tube (Eustachian tube) • Otitis Media – Connects the middle ear to the nasopharynx • Equalizes pressure – Opens during swallowing and yawning Middle Ear Middle Ear • Contains auditory ossicles (bones) • Sound waves cause tympanic membrane to vibrate – Malleus • Ossicles help transmit vibrations into the inner ear – Incus – Reduce the area -

Unit 13 Eye and Ear.Pdf
1 BIOL 2210L Unit 13: Eye and Ear Authors: Terri Koontz and Brandy Johnson, CNM Biology Department Creative Commons Attribution-NonCommercial 4.0 International License Terms to Know for Unit 13 Eye Neural parts of the eye Additional Instructor Terms Accessory structures of the eye Retina Conjunctiva Photoreceptors Lacrimal gland Optic disc Lacrimal sac Macula lutea Extrinsic eye muscles Fovea centralis Optic nerve Wall layers of the eye Fibrous tunic Ear Sclera Outer ear Cornea Pinna Vascular tunic External auditory canal Choroid Ciliary body Middle ear Ciliary muscles Tympanic membrane Iris Auditory ossicles Pupil Malleus Sensory tunic Incus Stapes Optics of the eye Auditory tube Lens Suspensory ligaments Inner ear Anterior segment of eye Semicircular canals Anterior chamber Crista ampullaris Posterior chamber Vestibule Aqueous humor Cochlea Canal of Schlemm Organ of Corti Posterior segment of eye Vitreous humor Learning Objectives (modified from HAPS learning outcomes) 1. Gross & microscopic anatomy of the eye a. Identify the accessory eye structures, the tunics, the optical components and the neural components of the eye. 2. Roles of specific tissues of the eye in vision 2 a. Describe the functions of the accessory structures of the eye. b. Trace the path of light as it passes through the eye to the retina and the path of nerve impulses from the retina to various parts of the brain. c. Describe the structure of the retina and the cells that compose it. d. Compare and contrast the function of rods and cones in vision. 3. General gross & microscopic anatomy of the hearing & accessory structures of the ear a. -

Tympanic Membrane (Membrana Tympanica, Myrinx)
Auditory and vestibular system Auris, is = Us, oton Auditory and vestibular system • external ear (auris externa) • middle ear (auris media) • internal ear (auris interna) = organum vestibulo- cochleare External ear (Auris externa) • auricle (auricula, pinna) – elastic cartilage • external acoustic meatus (meatus acusticus externus) • tympanic membrane (membrana tympanica, myrinx) • helix Auricle – crus, spina, cauda – (tuberculum auriculare Darwini, apex auriculae) • antihelix – crura, fossa triangularis • scapha • concha auriculae – cymba, cavitas • tragus • antitragus • incisura intertragica • lobulus auriculae posterior surface = negative image of the anterior one ligaments: lig. auriculare ant., sup., post. muscles – innervation: n. facialis • extrinsic muscles = facial muscles – mm. auriculares (ant., sup., post.) – m. temporoparietalis • intrinsic muscles: rudimentary – m. tragicus + antitragicus – m. helicis major+minor – m. obliquus + transversus auriculae, m. pyramidalis auriculae cartilage: cartilago auriculae - elastic skin: dorsally more loosen, ventrally firmly fixed to perichondrium - othematoma Auricle – supply • arteries: a. temporalis superficialis → rr. auriculares ant. a. carotis externa → a. auricularis post. • veins: v. jugularis ext. • lymph: nn.ll. parotidei, mastoidei • nerves: sensory – nn. auriculares ant. from n. auriculotemporalis (ventrocranial 2/3) – r. auricularis n. X. (concha) – n. occipitalis minor (dosrocranial) – n. auricularis magnus (cudal) motor: n. VII. External acoustic meatus (meatus acusticus -
Hair Cell Regeneration in the Avian Vestibular Epithelium
EXPERIMENTAL NEUROLOGY 115,2-6 (1992) Hair Cell Regeneration in the Avian Vestibular Epithelium PEDRO WEISLEDER AND EDWIN W RUBEL Hearing Development Laboratories, RL-30, University of Washington, Seattle, Washington 98195 the epithelium. These cells are cuboidal in morphology, Research conducted in the past 4 years has shown rest on the basement membrane, and extend to the free that the avian vestibular system retains the capacity to surface of the epithelium. On the lumenal surface of the generate hair cells postnatally. In the present paper we supporting cells, several microvilli and a modified short review information on postnatal proliferation and dif- kinocilium can be seen. ferentiation of hair cells in the avian vestibular system. In addition, we present preliminary accounts of recent experiments regarding regeneration of vestibular hair ONGOING PROLIFERATION IN THE cells following aminoglycoside toxicity. The overall VESTIBULAR EPITHELIUM consensus is that the avian vestibular system is able to regenerate hair cells, both on an ongoing basis and after Jorgensen and Mathiesen (5) reported that a low, on- damage. 0 1992 Academic Press, Inc. going level of cell proliferation could be detected in the vestibular system of adult budgerigars (Australian parrots). Analysis of tissue from tritiated thymidine-in- INTRODUCTION jetted animals revealed labeled cells in the sensory epi- thelium of both ampullary and otolithic organs. Labeled Until recently, it has been believed that proliferation nuclei of Type II hair cells and supporting cells were and differentiation of hair cells in the inner ear of mam- seen throughout the parenchyma, indicating that newly mals and birds occurred only during prenatal and early formed sensory and supporting cells were added to the postnatal development (12).