The Degree of Altriciality and Performance in a Cognitive Task Show Correlated Evolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Altriciality and the Evolution of Toe Orientation in Birds
Evol Biol DOI 10.1007/s11692-015-9334-7 SYNTHESIS PAPER Altriciality and the Evolution of Toe Orientation in Birds 1 1 1 Joa˜o Francisco Botelho • Daniel Smith-Paredes • Alexander O. Vargas Received: 3 November 2014 / Accepted: 18 June 2015 Ó Springer Science+Business Media New York 2015 Abstract Specialized morphologies of bird feet have trees, to swim under and above the water surface, to hunt and evolved several times independently as different groups have fish, and to walk in the mud and over aquatic vegetation, become zygodactyl, semi-zygodactyl, heterodactyl, pam- among other abilities. Toe orientations in the foot can be prodactyl or syndactyl. Birds have also convergently described in six main types: Anisodactyl feet have digit II evolved similar modes of development, in a spectrum that (dII), digit III (dIII) and digit IV (dIV) pointing forward and goes from precocial to altricial. Using the new context pro- digit I (dI) pointing backward. From the basal anisodactyl vided by recent molecular phylogenies, we compared the condition four feet types have arisen by modifications in the evolution of foot morphology and modes of development orientation of digits. Zygodactyl feet have dI and dIV ori- among extant avian families. Variations in the arrangement ented backward and dII and dIII oriented forward, a condi- of toes with respect to the anisodactyl ancestral condition tion similar to heterodactyl feet, which have dI and dII have occurred only in altricial groups. Those groups repre- oriented backward and dIII and dIV oriented forward. Semi- sent four independent events of super-altriciality and many zygodactyl birds can assume a facultative zygodactyl or independent transformations of toe arrangements (at least almost zygodactyl orientation. -

Birds of the East Texas Baptist University Campus with Birds Observed Off-Campus During BIOL3400 Field Course
Birds of the East Texas Baptist University Campus with birds observed off-campus during BIOL3400 Field course Photo Credit: Talton Cooper Species Descriptions and Photos by students of BIOL3400 Edited by Troy A. Ladine Photo Credit: Kenneth Anding Links to Tables, Figures, and Species accounts for birds observed during May-term course or winter bird counts. Figure 1. Location of Environmental Studies Area Table. 1. Number of species and number of days observing birds during the field course from 2005 to 2016 and annual statistics. Table 2. Compilation of species observed during May 2005 - 2016 on campus and off-campus. Table 3. Number of days, by year, species have been observed on the campus of ETBU. Table 4. Number of days, by year, species have been observed during the off-campus trips. Table 5. Number of days, by year, species have been observed during a winter count of birds on the Environmental Studies Area of ETBU. Table 6. Species observed from 1 September to 1 October 2009 on the Environmental Studies Area of ETBU. Alphabetical Listing of Birds with authors of accounts and photographers . A Acadian Flycatcher B Anhinga B Belted Kingfisher Alder Flycatcher Bald Eagle Travis W. Sammons American Bittern Shane Kelehan Bewick's Wren Lynlea Hansen Rusty Collier Black Phoebe American Coot Leslie Fletcher Black-throated Blue Warbler Jordan Bartlett Jovana Nieto Jacob Stone American Crow Baltimore Oriole Black Vulture Zane Gruznina Pete Fitzsimmons Jeremy Alexander Darius Roberts George Plumlee Blair Brown Rachel Hastie Janae Wineland Brent Lewis American Goldfinch Barn Swallow Keely Schlabs Kathleen Santanello Katy Gifford Black-and-white Warbler Matthew Armendarez Jordan Brewer Sheridan A. -

A Comprehensive Multilocus Assessment of Sparrow (Aves: Passerellidae) Relationships ⇑ John Klicka A, , F
Molecular Phylogenetics and Evolution 77 (2014) 177–182 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Short Communication A comprehensive multilocus assessment of sparrow (Aves: Passerellidae) relationships ⇑ John Klicka a, , F. Keith Barker b,c, Kevin J. Burns d, Scott M. Lanyon b, Irby J. Lovette e, Jaime A. Chaves f,g, Robert W. Bryson Jr. a a Department of Biology and Burke Museum of Natural History and Culture, University of Washington, Box 353010, Seattle, WA 98195-3010, USA b Department of Ecology, Evolution, and Behavior, University of Minnesota, 100 Ecology Building, 1987 Upper Buford Circle, St. Paul, MN 55108, USA c Bell Museum of Natural History, University of Minnesota, 100 Ecology Building, 1987 Upper Buford Circle, St. Paul, MN 55108, USA d Department of Biology, San Diego State University, San Diego, CA 92182, USA e Fuller Evolutionary Biology Program, Cornell Lab of Ornithology, Cornell University, 159 Sapsucker Woods Road, Ithaca, NY 14950, USA f Department of Biology, University of Miami, 1301 Memorial Drive, Coral Gables, FL 33146, USA g Universidad San Francisco de Quito, USFQ, Colegio de Ciencias Biológicas y Ambientales, y Extensión Galápagos, Campus Cumbayá, Casilla Postal 17-1200-841, Quito, Ecuador article info abstract Article history: The New World sparrows (Emberizidae) are among the best known of songbird groups and have long- Received 6 November 2013 been recognized as one of the prominent components of the New World nine-primaried oscine assem- Revised 16 April 2014 blage. Despite receiving much attention from taxonomists over the years, and only recently using molec- Accepted 21 April 2014 ular methods, was a ‘‘core’’ sparrow clade established allowing the reconstruction of a phylogenetic Available online 30 April 2014 hypothesis that includes the full sampling of sparrow species diversity. -

North Texas Winter Sparrows Cheat Sheet *Indicates Diagnostic Feature(S)
North Texas Winter Sparrows Cheat Sheet *Indicates diagnostic feature(s) Genus Spizella 1) American Tree Sparrow - “Average” bird 6.25 inches +/- - Long notched tail - *Thin streak-like rufous patch on the side of the breast - Two white wing bars, upper bar distinctly shorter - *Distinct dark spot on otherwise clear greyish breast - Rufous crown and eye-line on otherwise grey face - *Upper mandible noticeable darker than lower - Prefers brushy edges o Similar species; field sparrow (smaller and paler plumage) 2) Field Sparrow - Smaller bird 5.5 inches +/- “plump” - No streaking on pale grey breast some individuals may have faint reddish flanks - Long narrow tail - A lot of grey on the face, some individuals may have a faint darker grey or reddish eye streak - Two white wing bars, upper bar distinctly shorter - Rufous crown - White eye ring - *Pink bill; in combination with white eye ring and un-streaked breast - Open grasslands and woodland edges o Similar species; American Tree Sparrow (noticeably larger) 3) Chipping Sparrow - Smaller bird 5.5 inches - No streaking on plain grey breast and under parts - Rufous crown sometimes faintly streaked - Two white wing bars, upper bar distinctly shorter - *Dark eye-line coupled with lighter “eyebrow” stripe and rufous crown - Grey rump - Pink bill and legs - Common all over the place in winter o Similar species; Clay-colored Sparrow, a rare migrant Genus Ammodramus 4) Grasshopper Sparrow – a rare winter visitor - Small bird 5 inches - Thick “heavier” bill - Pointed, short tail compared to body -

Melospiza Melodia) Along an Urban Gradient Andrew Bartlow1, Katie White1, and Jeffrey A
The Relationship Between Hemosporidian Parasites and Song Sparrows (Melospiza melodia) Along an Urban Gradient Andrew Bartlow1, Katie White1, and Jeffrey A. Stratford1,2 1Department of Biology, Wilkes University, Wilkes-Barre, PA 2 [email protected] ABSTRACT Humans turn natural landscapes into complex mosaics, which include A. B. agriculture, urban centers, suburbs, and natural remnants. Consequently, interactions between organisms are likely to be altered including parasite- host interactions. Here, we investigate the relationship between song 50 The results show that the probability of sparrows (Melospiza melodia) and several hematozoan parasites in a hemosporidian parasite infection increases with complex landscape within and around Wilkes-Barre, PA. Fifty-four 40 35 increasing forest cover at a local scale (90m around a sparrows were captured in various habitats from urban centers to 28 sample point) and increasing urban cover at a larger relatively rural sites. From each bird, a blood smear was made and 30 scale (210m around a sample point). examined for 15 minutes (30 minutes total) by two observers noting presence/absence of hematozoans. Capture sites served as centers of 20 circular buffers from which we extracted the proportion of urban and 10 10 forest cover. Buffers had diameters of 90, 210 and 990m around each 10 RESULTS point. We created a number of logistic models with presence/absence as C. D. of Infections Percent • Plasmodium is the most frequently detected parasite (Fig. 3) the response variable and arcsine-square-root transformed landscape 0 • The top model includes small scale forest and medium scale variables as the explanatory variables. Using Akaiki Information Criterion Leucocytozoon Plasmodium Haemoproteus Other urban cover (Table 1) (AIC) to rank models, the top model included percent forest cover in 90m • Forest on a small scale within a slightly larger urban landscape buffers and percent urban in 210m buffers. -

Predictors of Juvenile Survival in Birds
Ornithological Monographs Volume (2013), No. 78, 1–55 © The American Ornithologists’ Union, 2013. Printed in USA. PREDICTORS OF JUVENILE SURVIVAL IN BIRDS TERRI J. MANESS1,2,3 AND DAVID J. ANDERSON2 1School of Biological Sciences, Louisiana Tech University, Ruston, Louisiana 71272, USA; and 2Department of Biology, Wake Forest University, Winston-Salem, North Carolina 27106, USA ABSTRACT.—The survival probability of birds during the juvenile period, between the end of parental care and adulthood, is highly variable and has a major effect on population dynamics and parental fitness. As such, a large number of studies have attempted to evaluate potential predictors of juvenile survival in birds, especially predictors related to parental care. Lack’s hypothesis linking body reserves accumulated from parental care to the survival of naive juveniles has organized much of this research, but various other predictors have also been investigated and received some support. We reviewed the literature in this area and identified a variety of methodological problems that obscure interpretation of the body of results. Most studies adopted statistical techniques that missed the opportunities to (1) evaluate the relative importance of several predictors, (2) control the confounding effect of correlation among predictor variables, and (3) exploit the information content of collinearity by evaluating indirect (via correlation) as well as direct effects of potential predictors on juvenile survival. Ultimately, we concluded that too few reliable studies exist to allow robust evaluations of any hypothesis regarding juvenile survival in birds. We used path analysis to test potential predictors of juvenile survival of 2,631 offspring from seven annual cohorts of a seabird, the Nazca Booby (Sula granti). -

Updating the Distribution of the Sierra Madre Sparrow Xenospiza Baileyi Across Central Mexico: Historical Records, New Localities, and Conservation Perspectives
VOLUME 15, ISSUE 1, ARTICLE 15 Ortega-Álvarez, R., R. Calderón-Parra, U. Martínez Molina, F. Martínez Molina, G. Martínez Molina, Y. Martínez Molina, A. Martínez Villagrán, J. Martínez Freire, R. Vásquez Robles, D. García Loaeza, J. Martínez García, S. García Loaeza, N. I. Garduño López, and L. A. Sánchez-González. 2020. Updating the distribution of the Sierra Madre Sparrow Xenospiza baileyi across central Mexico: historical records, new localities, and conservation perspectives. Avian Conservation and Ecology 15(1):15. https://doi.org/10.5751/ACE-01573-150115 Copyright © 2020 by the author(s). Published here under license by the Resilience Alliance. Research Paper Updating the distribution of the Sierra Madre Sparrow Xenospiza baileyi across central Mexico: historical records, new localities, and conservation perspectives Rubén Ortega-Álvarez 1, Rafael Calderón-Parra 2, Ulises Martínez Molina 2, Fredy Martínez Molina 2, Gabriel Martínez Molina 2, Yuridia Martínez Molina 2, Agustín Martínez Villagrán 2, Josué Martínez Freire 2, Rocío Vásquez Robles 2, Delfino García Loaeza 2, Jaciel Martínez García 2, Sarai García Loaeza 2, Nancy Isabel Garduño López 2 and Luis A. Sánchez-González 1 1Museo de Zoología "Alfonso L. Herrera," Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad de México, México, 2Brigada de Monitoreo Biológico Milpa Alta, San Pablo Oztotepec, Milpa Alta, Ciudad de México, México ABSTRACT. The Sierra Madre Sparrow (Xenospiza baileyi) is an endangered species microendemic to subalpine grasslands of central and northwestern Mexico. Given that land use change is highly dynamic in central Mexico, there is a need for a continuous monitoring of the distribution and conservation status of the species across the region. -
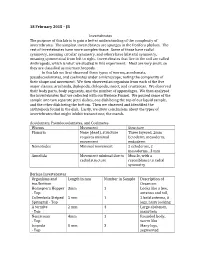
18 February 2015 – JB Invertebrates the Purpose of This Lab Is to Gain
18 February 2015 – JB Invertebrates The purpose of this lab is to gain a better understanding of the complexity of invertebrates. The simplest invertebrates are sponges in the Porifera phylum. The rest of invertebrates have more complex tissue. Some of those have radial symmetry, meaning circular symmetry, and others have bilateral symmetry, meaning symmetrical from left to right. Invertebrates that live in the soil are called Anthropods, which is what we studied in this experiment. Most are very small, so they are classified as microanthropods. In this lab we first observed three types of worms, acoelomats, pseudocoelomates, and coelmates under a microscope; noting the complexity of their shape and movement. We then observed an organism from each of the five major classes: arachnida, diplopoda, chilopoda, insect, and crustacean. We observed their body parts, body segments, and the number of appendages. We then analyzed the invertebrates that we collected with our Berlese Funnel. We poured some of the sample into two separate petri dishes, one dish being the top of our liquid sample, and the other dish being the bottom. Then we observed and identified the anthropods found in the dish. Lastly, we drew conclusions about the types of invertebrates that might inhibit transect one, the marsh. Acoelomats, Pseudocoelomates, and Coelmates Worms Movement Structure Planaria None (dead), structure Three layered, 2mm requires minimal Ectoderm, mesoderm, movement endoderm Nemotodes Minimal movement 2 ectoderms, 2 mesoderms , 3 mm Annelida Movement minimal due to Muscle, with a radial structure resemblance to radial symmetry Berlese Invertebrates Organisms and Length in mm Number in Sample Description of top/bottom Organism Homoptera Hopper 3mm 1 Looks like a bee, - Top antenna and tail, Collembola Striped 5 mm 1 2 bold antenna, 6 Springtail - Top legs, hairy looking A termite 2 mm 4 Large abdomen, - Top many legs Nematoma 4mm 1 Rounded body, - Top worm like Isopoda 8 mm 3 Many legs, - Top segmented The size range of the organisms observed was 2mm to 8mm. -

Illinois Birds: Volume 4 – Sparrows, Weaver Finches and Longspurs © 2013, Edges, Fence Rows, Thickets and Grain Fields
ILLINOIS BIRDS : Volume 4 SPARROWS, WEAVER FINCHES and LONGSPURS male Photo © Rob Curtis, The Early Birder female Photo © John Cassady Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Mary Kay Rubey Photo © Rob Curtis, The Early Birder American tree sparrow chipping sparrow field sparrow vesper sparrow eastern towhee Pipilo erythrophthalmus Spizella arborea Spizella passerina Spizella pusilla Pooecetes gramineus Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder lark sparrow savannah sparrow grasshopper sparrow Henslow’s sparrow fox sparrow song sparrow Chondestes grammacus Passerculus sandwichensis Ammodramus savannarum Ammodramus henslowii Passerella iliaca Melospiza melodia Photo © Brian Tang Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Photo © Rob Curtis, The Early Birder Lincoln’s sparrow swamp sparrow white-throated sparrow white-crowned sparrow dark-eyed junco Le Conte’s sparrow Melospiza lincolnii Melospiza georgiana Zonotrichia albicollis Zonotrichia leucophrys Junco hyemalis Ammodramus leconteii Photo © Brian Tang winter Photo © Rob Curtis, The Early Birder summer Photo © Rob Curtis, The Early Birder Photo © Mark Bowman winter Photo © Rob Curtis, The Early Birder summer Photo © Rob Curtis, The Early Birder Nelson’s sparrow -

The Platypus Is Not a Rodent: DNA Hybridization, Amniote Phylogeny and the Palimpsest Theory
The platypus is not a rodent: DNA hybridization, amniote phylogeny and the palimpsest theory John A. W. Kirsch1* and Gregory C. Mayer1,2 1The University of Wisconsin Zoological Museum, 250 North Mills Street, Madison,WI 53706, USA 2Department of Biological Sciences, University of Wisconsin-Parkside, Kenosha,WI 53141, USA We present DNA-hybridization data on 21 amniotes and two anurans showing that discrimination is obtained among most of these at the class and lower levels. Trees generated from these data largely agree with conventional views, for example in not associating birds and mammals. However, the sister relation- ships found here of the monotremes to marsupials, and of turtles to the alligator, are surprising results which are nonetheless consistent with the results of some other studies. The Marsupionta hypothesis of Gregory is reviewed, as are opinions about the placement of chelonians. Anatomical and reproductive data considered by Gregory do not unequivocally preclude a marsupial^monotreme special relationship, and there is other recent evidence for placing turtles within the Diapsida. We conclude that the evidential meaning of the molecular data is as shown in the trees, but that the topologies may be in£uenced by a base- compositional bias producing a seemingly slow evolutionary rate in monotremes, or by algorithmic artefacts (in the case of turtles as well). Keywords: Chelonia; Crocodilia; Eutheria; Marsupialia; Marsupionta; molecular evolution `Unyielding factualists have so set the style in taxonomy relationships continue to be debated, `everyone knows' and morphology that, if their assertions were accepted, that the latter result cannot be correct. hardly any known type of animal could possibly have The orthodox view of the phylogeny of mammals been derived even from any known past group.' regards monotremes (the living `Prototheria') as the Gregory (1947, p. -

Mammalian Organogenesis in Deep Time: Tools for Teaching and Outreach Marcelo R
Sánchez‑Villagra and Werneburg Evo Edu Outreach (2016) 9:11 DOI 10.1186/s12052-016-0062-y REVIEW Open Access Mammalian organogenesis in deep time: tools for teaching and outreach Marcelo R. Sánchez‑Villagra1 and Ingmar Werneburg1,2,3,4* Abstract Mammals constitute a rich subject of study on evolution and development and provide model organisms for experi‑ mental investigations. They can serve to illustrate how ontogeny and phylogeny can be studied together and how the reconstruction of ancestors of our own evolutionary lineage can be approached. Likewise, mammals can be used to promote ’tree thinking’ and can provide an organismal appreciation of evolutionary changes. This subject is suitable for the classroom and to the public at large given the interest and familiarity of people with mammals and their closest relatives. We present a simple exercise in which embryonic development is presented as a transforma‑ tive process that can be observed, compared, and analyzed. In addition, we provide and discuss a freely available animation on organogenesis and life history evolution in mammals. An evolutionary tree can be the best tool to order and understand those transformations for different species. A simple exercise introduces the subject of changes in developmental timing or heterochrony and its importance in evolution. The developmental perspective is relevant in teaching and outreach efforts for the understanding of evolutionary theory today. Keywords: Development, Ontogeny, Embryology, Phylogeny, Heterochrony, Recapitulation, Placentalia, Human Background (Gilbert 2013), followed by the growth process. In pla- Mammals are a diverse group in which to examine devel- cental mammals, organogenesis takes place mostly in the opment and evolution, and besides the mouse and the rat uterus, whereas in monotremes and marsupials a very used in biomedical research, provide subjects based on immature hatchling or newborn, respectively, develops which experimental (Harjunmaa et al. -

An Efficient Method of Capturing Painted Buntings and Other Small Granivorous Passerines
An Efficient Method of Capturing Painted Buntings and Other Small Granivorous Passerines Paul W. Sykes, Jr. USGS Patuxent Wildlife Research Center populations, among them research on annual Warnell School of Forestry and Natural Res. survival of the eastern population (Sykes unpub The University of Georgia lished data). As part of this research, captures of Athens, GA 30602 large numbers of buntings are necessary for [email protected] subsequent study. I developed a methodology to achieve this goal, which is herein described. ABSTRACT METHODS To study survival in the eastern breeding population of the Painted Bunting (Passerina ciris), I developed Painted Buntings are attracted to bird feeders a technique to capture a large sample of buntings provisioned with white prose millet (Panicum verg1) for color marking with leg-bands. This involved the creating a focal point where birds can be trapped. use of bird feeders and an array of three short mist Twenty study areas were selected randomly within nets located at 40 sites in four states, each site the breeding range of the eastern population, five meeting five specific criteria. In five years of mist each in Florida, Georgia, South Carolina, and North netting (1999-2003), 4174 captures (including Carolina. Within each study area, two sites were recaptures) of Painted Buntings were made in 3393 selected; thus there were 40 sites, ten per state. net-hours or 123 captures per 100 net-hours. The For practical reasons, each study site had to meet technique proved to be effective and efficient, and the following five criteria: may have broad application for capturing large (1) had a high density of buntings present, numbers of small granivorous passerines.