The Vertical Distribution of Fish on Two Offshore Oil Platforms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Zootaxa 457: 1–8 (2004) ISSN 1175-5326 (Print Edition) ZOOTAXA 457 Copyright © 2004 Magnolia Press ISSN 1175-5334 (Online Edition)
Zootaxa 457: 1–8 (2004) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA 457 Copyright © 2004 Magnolia Press ISSN 1175-5334 (online edition) Stephanostomum tantabiddii n. sp. (Digenea: Acanthocolpidae) from Carangoides fulvoguttatus (Forsskål, 1775) (Perciformes: Carangidae) from Ningaloo Reef, Western Australia RODNEY A. BRAY1 & THOMAS H. CRIBB2 1Department of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD, UK, email: [email protected] 2Centre for Marine Studies and Department of Microbiology and Parasitology, The University of Queensland, Brisbane, Queensland 4074, Australia, email: [email protected] Abstract A new species, Stephanostomum tantabiddii n. sp., is described from the yellowspotted trevally Carangoides fulvoguttatus from Ningaloo Reef, Western Australia. It has 38–45 circum-oral spines and the vitellarium reaches to no less than 17% of the hindbody length from the ventral sucker. It differs from other species of Stephanostomum with these characteristics by various combinations of the ventral hiatus of the circum-oral spine rows, the relatively long pars prostatica and short ejacu- latory duct, the elongate body and the wide gaps between the gonads. Key words: Stephanostomum tantabiddii n. sp., Acanthocolpidae, Digenea, Carangoides fulvogut- tatus, Carangidae, Perciformes, Ningaloo Coral Reef, Western Australia Introduction Members of the large genus Stephanostomum Looss, 1899 parasitise a wide variety of teleost hosts, but a significant proportion of them are found in fishes of the perciform fam- ily Carangidae. This family includes many large pelagic piscivores. The second intermedi- ate hosts of Stephanostomum are almost always found to be fishes and, as piscivores, carangids are therefore particularly susceptible to infection (Pérez-Urbiola & Martínez- Díaz, 2001). -

MARKET FISHES of INDONESIA Market Fishes
MARKET FISHES OF INDONESIA market fishes Market fishes indonesiaof of Indonesia 3 This bilingual, full-colour identification William T. White guide is the result of a joint collaborative 3 Peter R. Last project between Indonesia and Australia 3 Dharmadi and is an essential reference for fish 3 Ria Faizah scientists, fisheries officers, fishers, 3 Umi Chodrijah consumers and enthusiasts. 3 Budi Iskandar Prisantoso This is the first detailed guide to the bony 3 John J. Pogonoski fish species that are caught and marketed 3 Melody Puckridge in Indonesia. The bilingual layout contains information on identifying features, size, 3 Stephen J.M. Blaber distribution and habitat of 873 bony fish species recorded during intensive surveys of fish landing sites and markets. 155 market fishes indonesiaof jenis-jenis ikan indonesiadi 3 William T. White 3 Peter R. Last 3 Dharmadi 3 Ria Faizah 3 Umi Chodrijah 3 Budi Iskandar Prisantoso 3 John J. Pogonoski 3 Melody Puckridge 3 Stephen J.M. Blaber The Australian Centre for International Agricultural Research (ACIAR) was established in June 1982 by an Act of the Australian Parliament. ACIAR operates as part of Australia’s international development cooperation program, with a mission to achieve more productive and sustainable agricultural systems, for the benefit of developing countries and Australia. It commissions collaborative research between Australian and developing-country researchers in areas where Australia has special research competence. It also administers Australia’s contribution to the International Agricultural Research Centres. Where trade names are used, this constitutes neither endorsement of nor discrimination against any product by ACIAR. ACIAR MONOGRAPH SERIES This series contains the results of original research supported by ACIAR, or material deemed relevant to ACIAR’s research and development objectives. -

Reksten Et Al.2020 Nutrient Composition.Pdf
Journal of Food Composition and Analysis 91 (2020) 103508 Contents lists available at ScienceDirect Journal of Food Composition and Analysis journal homepage: www.elsevier.com/locate/jfca Original Research Article Nutrient composition of 19 fish species from Sri Lanka and potential T contribution to food and nutrition security Amalie Moxness Rekstena,*, Thiruchenduran Somasundaramb, Marian Kjellevolda, Anna Nordhagena, Annbjørg Bøkevolla, Lauren Michelle Pincusc, Abu Ansar Md. Rizwand, Al Mamune, Shakuntala Haraksingh Thilstedc, Thaung Htutf, Inger Aakrea a Institute of Marine Research, P.O. Box 2029 Nordnes, 5817 Bergen, Norway b Institute of Postharvest Technology, National Aquatic Resources Research and Development Agency, Colombo, Sri Lanka c WorldFish, Jalan Batu Muang, Batu Muang, Bayan Lepas, 11960 Penang, Malaysia d Health and Nutrition, Social Assistance and Rehabilitation for the Physically Vulnerable (SARPV), Cox’s Bazar, Bangladesh e Marine Fisheries Survey Management Unit, Department of Fisheries, Bangladesh f Wildlife Conservation Society-Myanmar Program, P.O. Box Kamayut, 11041 Yangon, Myanmar ARTICLE INFO ABSTRACT Keywords: Fish is an important part of the Sri Lankan diet. However, existing data on the nutrient composition of fish in Sri Food composition Lanka is highly outdated and limited. The aim of this study was to report the nutrient composition of commonly Fish consumed marine fish species in Sri Lanka and assess the potential contribution of selected key nutrients infish Marine to recommended nutrient intakes (RNI). Fish were sampled during a survey with research vessel Dr. Fridtjof Food analysis Nansen around Sri Lanka. Species were categorised as either small (< 25 cm, n = 12) or large (> 25 cm, n = 7), Sri Lanka and three composite samples from each species were analysed using accredited methods. -

Training Manual Series No.15/2018
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by CMFRI Digital Repository DBTR-H D Indian Council of Agricultural Research Ministry of Science and Technology Central Marine Fisheries Research Institute Department of Biotechnology CMFRI Training Manual Series No.15/2018 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual This is a limited edition of the CMFRI Training Manual provided to participants of the “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals” organized by the Marine Biotechnology Division of Central Marine Fisheries Research Institute (CMFRI), from 2nd February 2015 - 31st March 2018. Principal Investigator Dr. P. Vijayagopal Compiled & Edited by Dr. P. Vijayagopal Dr. Reynold Peter Assisted by Aditya Prabhakar Swetha Dhamodharan P V ISBN 978-93-82263-24-1 CMFRI Training Manual Series No.15/2018 Published by Dr A Gopalakrishnan Director, Central Marine Fisheries Research Institute (ICAR-CMFRI) Central Marine Fisheries Research Institute PB.No:1603, Ernakulam North P.O, Kochi-682018, India. 2 Foreword Central Marine Fisheries Research Institute (CMFRI), Kochi along with CIFE, Mumbai and CIFA, Bhubaneswar within the Indian Council of Agricultural Research (ICAR) and Department of Biotechnology of Government of India organized a series of training programs entitled “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals”. -

Nematode Parasites of Four Species of Carangoides (Osteichthyes: Carangidae) in New Caledonian Waters, with a Description of Philometra Dispar N
Nematode parasites of four species of Carangoides (Osteichthyes: Carangidae) in New Caledonian waters, with a description of Philometra dispar n. sp. (Philometridae) František Moravec, Delphine Gey, Jean-Lou Justine To cite this version: František Moravec, Delphine Gey, Jean-Lou Justine. Nematode parasites of four species of Carangoides (Osteichthyes: Carangidae) in New Caledonian waters, with a description of Philometra dispar n. sp. (Philometridae). Parasite, EDP Sciences, 2016, 23, pp.40. 10.1051/parasite/2016049. hal-01399891 HAL Id: hal-01399891 https://hal.sorbonne-universite.fr/hal-01399891 Submitted on 21 Nov 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License Parasite 2016, 23,40 Ó F. Moravec et al., published by EDP Sciences, 2016 DOI: 10.1051/parasite/2016049 urn:lsid:zoobank.org:pub:C2F6A05A-66AC-4ED1-82D7-F503BD34A943 Available online at: www.parasite-journal.org RESEARCH ARTICLE OPEN ACCESS Nematode parasites of four species of Carangoides (Osteichthyes: Carangidae) -

Marine Ecology Progress Series 530:223
The following supplement accompanies the article Economic incentives and overfishing: a bioeconomic vulnerability index William W. L. Cheung*, U. Rashid Sumaila *Corresponding author: [email protected] Marine Ecology Progress Series 530: 223–232 (2015) Supplement Table S1. Country level discount rate used in the analysis Country/Territory Discount rate (%) Albania 13.4 Algeria 8.0 Amer Samoa 11.9 Andaman Is 10.0 Angola 35.0 Anguilla 10.0 Antigua Barb 10.9 Argentina 8.6 Aruba 11.3 Ascension Is 10.0 Australia 6.5 Azores Is 7.0 Bahamas 5.3 Bahrain 8.1 Baker Howland Is 7.0 Bangladesh 15.1 Barbados 9.7 Belgium 3.8 Belize 14.3 Benin 10.0 Bermuda 7.0 Bosnia Herzg 10.0 Bouvet Is 7.0 Br Ind Oc Tr 7.0 Br Virgin Is 10.0 Brazil 50.0 Brunei Darsm 10.0 Country/Territory Discount rate (%) Bulgaria 9.2 Cambodia 16.9 Cameroon 16.0 Canada 8.0 Canary Is 7.0 Cape Verde 12.3 Cayman Is 7.0 Channel Is 7.0 Chile 7.8 China Main 5.9 Christmas I. 10.0 Clipperton Is 7.0 Cocos Is 10.0 Colombia 14.3 Comoros 10.8 Congo Dem Rep 16.0 Congo Rep 16.0 Cook Is. 10.0 Costa Rica 19.9 Cote d'Ivoire 10.0 Croatia 10.0 Crozet Is 7.0 Cuba 10.0 Cyprus 6.8 Denmark 7.0 Desventuradas Is 10.0 Djibouti 11.2 Dominica 9.5 Dominican Rp 19.8 East Timor 10.0 Easter Is 10.0 Ecuador 9.4 Egypt 12.8 El Salvador 10.0 Eq Guinea 16.0 Eritrea 10.0 Estonia 10.0 Faeroe Is 7.0 Falkland Is 7.0 Fiji 6.2 Finland 7.0 Fr Guiana 10.0 Fr Moz Ch Is 10.0 Country/Territory Discount rate (%) Fr Polynesia 10.0 France 4.0 Gabon 16.0 Galapagos Is 10.0 Gambia 30.9 Gaza Strip 10.0 Georgia 20.3 Germany (Baltic) 7.0 Germany (North Sea) 7.0 Ghana 10.0 Gibraltar 7.0 Greece 7.0 Greenland 7.0 Grenada 9.9 Guadeloupe 10.0 Guam 7.0 Guatemala 12.9 Guinea 10.0 GuineaBissau 10.0 Guyana 14.6 Haiti 43.8 Heard Is 7.0 Honduras 17.6 Hong Kong 7.4 Iceland 17.3 India 11.7 Indonesia 16.0 Iran 15.0 Iraq 14.1 Ireland 2.7 Isle of Man 7.0 Israel 6.9 Italy 5.8 Jamaica 17.5 Jan Mayen 7.0 Japan (Pacific Coast) 10.0 Japan (Sea of Japan) 10.0 Jarvis Is 10.0 Johnston I. -

Nematode Parasites of Four Species of Carangoides (Osteichthyes: Carangidae) in New Caledonian Waters, with a Description of Philometra Dispar N
Parasite 2016, 23,40 Ó F. Moravec et al., published by EDP Sciences, 2016 DOI: 10.1051/parasite/2016049 urn:lsid:zoobank.org:pub:C2F6A05A-66AC-4ED1-82D7-F503BD34A943 Available online at: www.parasite-journal.org RESEARCH ARTICLE OPEN ACCESS Nematode parasites of four species of Carangoides (Osteichthyes: Carangidae) in New Caledonian waters, with a description of Philometra dispar n. sp. (Philometridae) František Moravec1,*, Delphine Gey2, and Jean-Lou Justine3 1 Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, Branišovská 31, 370 05 Cˇ eské Budeˇjovice, Czech Republic 2 Service de Systématique moléculaire, UMS 2700 CNRS, Muséum National d’Histoire Naturelle, Sorbonne Universités, CP 26, 43 rue Cuvier, 75231 Paris cedex 05, France 3 ISYEB, Institut Systématique, Évolution, Biodiversité, UMR7205 CNRS, EPHE, MNHN, UPMC, Muséum National d’Histoire Naturelle, Sorbonne Universités, CP51, 57 rue Cuvier, 75231 Paris cedex 05, France Received 10 August 2016, Accepted 28 August 2016, Published online 12 September 2016 Abstract – Parasitological examination of marine perciform fishes belonging to four species of Carangoides, i.e. C. chrysophrys, C. dinema, C. fulvoguttatus and C. hedlandensis (Carangidae), from off New Caledonia revealed the presence of nematodes. The identification of carangids was confirmed by barcoding of the COI gene. The eight nematode species found were: Capillariidae gen. sp. (females), Cucullanus bulbosus (Lane, 1916) (male and females), Hysterothylacium sp. third-stage larvae, Raphidascaris (Ichthyascaris) sp. (female and larvae), Terranova sp. third- stage larvae, Philometra dispar n. sp. (male), Camallanus carangis Olsen, 1954 (females) and Johnstonmawsonia sp. (female). The new species P. dispar from the abdominal cavity of C. -
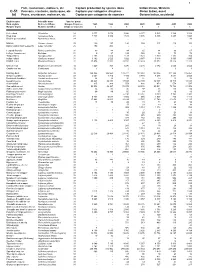
Fish, Crustaceans, Molluscs, Etc Capture
1 Fish, crustaceans, molluscs, etc Capture production by species items Indian Ocean, Western C-51 Poissons, crustacés, mollusques, etc Captures par catégories d'espèces Océan Indien, ouest (a) Peces, crustáceos, moluscos, etc Capturas por categorías de especies Océano Indico, occidental English name Scientific name Species group Nom anglais Nom scientifique Groupe d'espèces 1998 1999 2000 2001 2002 2003 2004 Nombre inglés Nombre científico Grupo de especies t t t t t t t Kelee shad Hilsa kelee 24 3 077 3 076 3 896 4 277 5 525 2 306 3 208 Hilsa shad Tenualosa ilisha 24 7 747 6 482 4 545 3 846 6 146 8 257 4 024 Bloch's gizzard shad Nematalosa nasus 24 ... ... ... ... ... ... 93 Milkfish Chanos chanos 25 139 137 125 154 117 113 101 Barramundi(=Giant seaperch) Lates calcarifer 25 196 204 - - - - - Leopard flounder Bothus pantherinus 31 90 84 84 65 84 96 67 Lefteye flounders nei Bothidae 31 9 27 129 125 88 73 78 Tonguefishes Cynoglossidae 31 2 158 2 066 2 137 1 923 1 989 1 383 984 Indian halibut Psettodes erumei 31 3 192 2 520 3 022 2 957 3 118 3 365 4 095 Flatfishes nei Pleuronectiformes 31 17 474 12 851 24 720 11 489 15 573 15 924 11 325 Unicorn cod Bregmaceros mcclellandi 32 1 449 743 1 470 2 435 2 372 2 643 2 624 Gadiformes nei Gadiformes 32 7 5 2 1 - 1 3 Bombay-duck Harpadon nehereus 33 144 865 146 663 133 221 141 082 100 366 101 936 116 452 Greater lizardfish Saurida tumbil 33 6 064 3 710 3 485 3 894 4 247 3 634 2 655 Brushtooth lizardfish Saurida undosquamis 33 34 20 30 32 20 11 18 Lizardfishes nei Synodontidae 33 20 221 20 871 17 894 12 433 12 447 10 715 13 833 Giant catfish Arius thalassinus 33 315 309 554 480 470 574 597 Sea catfishes nei Ariidae 33 99 932 98 467 78 375 89 308 89 941 78 788 81 972 Sabre squirrelfish Sargocentron spiniferum 33 34 53 46 97 96 135 114 Flathead grey mullet Mugil cephalus 33 69 89 60 34 37 40 46 Klunzinger's mullet Liza klunzingeri 33 .. -

Western Indian Ocean JOURNAL of Marine Science
Western Indian Ocean JOURNAL OF Marine Science Volume 18 | Issue 2 | Jul – Dec 2019 | ISSN: 0856-860X Chief Editor José Paula Western Indian Ocean JOURNAL OF Marine Science Chief Editor José Paula | Faculty of Sciences of University of Lisbon, Portugal Copy Editor Timothy Andrew Editorial Board Lena GIPPERTH Aviti MMOCHI Sweden Tanzania Serge ANDREFOUËT Johan GROENEVELD France Cosmas MUNGA South Africa Kenya Ranjeet BHAGOOLI Issufo HALO Mauritius South Africa/Mozambique Nyawira MUTHIGA Kenya Salomão BANDEIRA Christina HICKS Mozambique Australia/UK Brent NEWMAN Betsy Anne BEYMER-FARRIS Johnson KITHEKA South Africa USA/Norway Kenya Jan ROBINSON Jared BOSIRE Kassim KULINDWA Seycheles Kenya Tanzania Sérgio ROSENDO Atanásio BRITO Thierry LAVITRA Portugal Mozambique Madagascar Louis CELLIERS Blandina LUGENDO Melita SAMOILYS Kenya South Africa Tanzania Pascale CHABANET Joseph MAINA Max TROELL France Australia Sweden Published biannually Aims and scope: The Western Indian Ocean Journal of Marine Science provides an avenue for the wide dissem- ination of high quality research generated in the Western Indian Ocean (WIO) region, in particular on the sustainable use of coastal and marine resources. This is central to the goal of supporting and promoting sustainable coastal development in the region, as well as contributing to the global base of marine science. The journal publishes original research articles dealing with all aspects of marine science and coastal manage- ment. Topics include, but are not limited to: theoretical studies, oceanography, marine biology and ecology, fisheries, recovery and restoration processes, legal and institutional frameworks, and interactions/relationships between humans and the coastal and marine environment. In addition, Western Indian Ocean Journal of Marine Science features state-of-the-art review articles and short communications. -

SFA Technical Report ISSN 1659-7141
Seychelles artisanal fisheries statistics for 2007 Item Type Report Authors Assan, C.N.; Dorizo, J.L. Publisher Seychelles Fishing Authority Download date 01/10/2021 13:50:09 Item License http://creativecommons.org/licenses/by-nc/3.0/ Link to Item http://hdl.handle.net/1834/4973 SFA Technical Report ISSN 1659-7141 SFA/R&D/065 SEYCHELLES ARTISANAL FISHERIES STATISTICS FOR 2007 MARCH 2009 SEYCHELLES FISHING AUTHORITY Data presented in this report was collected by Staff of the Seychelles Fishing Authority (SFA) Fisheries Research Section. This report was compiled and edited by the Fisheries Statisticians, who wish to express their gratitude to all the staff of the statistics unit who gave their time to ensure the successful publication of this technical report. C.N. Assan, J. L. Dorizo S.F.A 2009 II SOMALIA SEYCHELLES EXCLUSIVE ECONOMIC ZONE 0° KENYA INDIAN OCEAN MAHÉ 5° S TANZANIA ALDABRA 10° S INDIAN COMORES OCEAN MOZAMBIQUE 15° S MADAGASCAR MAURITIUS 20° S REUNION 40° E 45° E 50° E 55° E 60° E 65° E 70° E ILE CURIEUSE THE SISTERS PRASLIN FÉLICITÉ COUSIN COUSINE MARIE ANNE NORTH ISLAND TROMPEUSE ROCKS LA DIGUE SILHOUETTE CHIMNEY MAMELLE ISLAND ROCKS 4° 30’ S BRISANS ROCKS RÉCIF SAINTE ANNE ISLAND FRIGATE VICTORIA ILE AUX CERFS ILE CONCEPTION ILE THÉRÈSE MAHÉ 55° 30’ E 56° E III EXPLANATION OF TABLES AND FIGURES The format of the present report is the same as that adopted from the Seychelles Artisanal Fisheries Statistics for 1994. Data are presented by boat gear type thus: Summary (all boat-gear data) Small boat fisheries (several boat-gear combinations) Whaler handline fishery Schooner line fishery Catch, effort and CPUE (all boat-gear type) Fish purchase and export Most information is self-explanatory but the following points should be noted: (a) Any discrepancy in the totals and the sum of the components is due to the rounding-up of data. -

Commercial and Bycatch Market Fishes Panay Island, Republic Of
Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines Nanarisari nga Isda nga Ginabaligya sa Merkado sa Isla sang Panay, Pilipinas Hiroyuki Motomura Ulysses B. Alama Nozomu Muto Ricardo P. Babaran Satoshi Ishikawa Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines 1 Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines Nanarisari nga Isda nga Ginabaligya sa Merkado sa Isla sang Panay, Pilipinas 2 H. Motomura · U. B. Alama · N. Muto · R. P. Babaran · S. Ishikawa (eds) For bibliographic purposes this book should be cited as follows: Motomura, H., U. B. Alama, N. Muto, R. P. Babaran, and S. Ishikawa (eds). 2017 (Jan.). Commercial and bycatch market fishes of Panay Island, Republic of the Philippines. The Kagoshima University Museum, Kagoshima, University of the Philippines Visayas, Iloilo, and Research Institute for Humanity and Nature, Kyoto. 246 pp, 911 figs Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines 3 Commercial and Bycatch Market Fishes ofPanay Island, Republic of the Philippines Edited by Hiroyuki Motomura, Ulysses B. Alama, Nozomu Muto, Ricardo P. Babaran, and Satoshi Ishikawa The Kagoshima University Museum, Japan University of the Philippines Visayas, Philippines Research Institute for Humanity and Nature, Japan 4 H. Motomura · U. B. Alama · N. Muto · R. P. Babaran · S. Ishikawa (eds) Copyright © 2017 by the Kagoshima University Museum, Kagoshima, University of the Philippines Visayas, Iloilo, and Research Institute for Humanity and Nature, Kyoto All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means without prior written permission from the publisher. -

(Per Kilo) Price Monitoring Report for the Month of June 2017 All Regions
REGIONAL MONTHLY PREVAILING RETAIL (PER KILO) PRICE MONITORING REPORT FOR THE MONTH OF JUNE 2017 ALL REGIONS CLASSIFICATION SCIENTIFIC NAME ENGLISH NAME LOCAL NAME CAR R-2 R-3 R-4A R-4B R-5 R-6 Prices in Philippine Peso A. FINFISHES A.1 Marine Ablennes hians Needle fish Balo 120-140 Acanthurus mata Elongate surgeonfish Labahita 200.00 Acanthurus nigrofuscus Surgeon Fish/Lavender Tang Labahita 200-240 Acanthurus sp. Surgeon Fish Labahita 60.00 Alepes djedaba Shrimp scad Salay salay 200.00 Amblygaster sirm Spotted sardinella Tamban/hawol-hawol 70.00 40-50 Bagudlong/Budburon/kala Atule mate Yellow tail scad pato/Salay-salay 100-120 80-160 Auxis rochei Bullet Tuna Aloy/Tulingan/perit/bodboron 120.00 90-180 120.00 Auxis rochei Bullet tuna BIG 60-80 Auxis rochei Bullet tuna SMALL 80.00 Tulingan/turingan/pidlaya Auxis thazard Frigate tuna n/Tangigi/mangko 160.00 130.00 120.00 70-140 120-140 Caesio caerulaurea Blue and Gold fusilier Dalagang bukid 240.00 130.00 Caesio cuning Redbelly yellowtail fusilier Dalagang bukid (lapad) 220-240 260.00 200.00 120-160 Caesio erythrogaster Yellow-tailed fusilier Dalagang Bukid 380.00 100.00 Caesio spp.; Pterocaesio spp. Fusilier Dalagang Bukid 150-260 Carangoides ferdau Blue Trevally Talakitok 250.00 140.00 Carangoides fulvoguttatus Yellowspotted trevally Talakitok/mansa 250.00 Carangoides malabaricus Malabar trevally Lison 100-240 Carangoides uii Trevally Talakitok 250.00 Caranx ignobilis Giant trevally SLICED 250.00 Caranx sexfasciatus Big Eye Trevally Talakitok/samin samin 350.00 320.00 Cephalopholis argus