New Primate Genus from the Miocene of Argentina
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

1 CURRICULUM VITAE E. Christopher Kirk Department of Anthropology Phone: (512) 471-0056 2201 Speedway Stop C3200 Fax: (512) 47
CURRICULUM VITAE E. Christopher Kirk Department of Anthropology Phone: (512) 471-0056 2201 Speedway Stop C3200 Fax: (512) 471-6535 University of Texas at Austin [email protected] Austin, Texas 78712 PROFESSIONAL APPOINTMENTS 2016-Present University Distinguished Teaching Professor, University of Texas at Austin 2015-Present Professor, Department of Anthropology, University of Texas at Austin 2014-Present Research Associate, Vertebrate Paleontology Laboratory, Jackson School Museum of Earth History, University of Texas at Austin 2009-2015 Associate Professor, Department of Anthropology, University of Texas at Austin 2005-2014 Assistant Research Professor, Texas Memorial Museum, University of Texas at Austin 2003-2009 Assistant Professor, Department of Anthropology, University of Texas at Austin 2000-2003 Instructor, Department of Biological Anthropology and Anatomy, Duke University EDUCATION Ph.D. 2003 Biological Anthropology and Anatomy: Duke University B.A. 1995 Anthropology: University of Texas at Austin PROFESSIONAL AND HONOR SOCIETY MEMBERSHIPS American Association of Physical Anthropology Society of Vertebrate Paleontology Phi Beta Kappa HONORS & AWARDS 2016 Member, Academy of Distinguished Teachers – Recognizes tenured faculty members who have made sustained and significant contributions to education, particularly at the undergraduate level; Nominated by the Department of Anthropology and the College of Liberal Arts 2016 Billy Carr Distinguished Teaching Fellowship – Awarded in recognition of my efforts to include students -

Boletín Antártico Chileno, Edición Especial
EL CONTINENTE DONDE EMPIEZA EL FUTURO Índice DIRECTOR Y REPRESENTANTE LEGAL Presentación José Retamales Espinoza 5 Prólogo 7 EDITOR Reiner Canales La relación entre Sudamérica y la Antártica 11 (E-mail: [email protected]) El pasado de la Antártica… ¿una incógnita develada? 13 COMITÉ EDITORIAL Marcelo Leppe Cartes Edgardo Vega Separación de la fauna marina antártica y sudamericana desde una aproximación molecular 21 Marcelo González Elie Poulin, Claudio González-Wevar, Mathias Hüne, Angie Díaz Marcelo Leppe Conexiones geológicas entre Sudamérica y Antártica 27 Francisco Hervé Allamand DIRECCIÓN DE ARTE Pablo Ruiz Teneb Adaptaciones al Medio Antártico y Biorrecursos 33 El interés por esudiar la biodiversidad microbiológica en la Antártica 35 DISEÑO / DIAGRAMACIÓN Jenny M. Blamey Oscar Giordano / Menssage Producciones El capital celular y molecular para vivir a temperaturas bajo cero en aguas antárticas 39 Víctor Bugueño / Menssage Producciones Marcelo González Aravena Las esrategias de las plantas antárticas para sobrevivir en ambientes extremos y su rol como biorrecursos 45 PORTADA Marco A. Molina-Montenegro, Ian S. Acuña-Rodríguez, Jorge Gallardo-Cerda, Rasme Hereme y Crisian Torres-Díaz EL CONTINENTE Arnaldo Gómez / Menssage Producciones DONDE EMPIEZA EL FUTURO Biodiversidad 53 A más de medio siglo IMPRESIÓN de la fundación del Menssage Producciones Las aves marinas de la península Antártica 55 Instituto Antártico Chileno Santiago de Chile Javier A. Arata El desafío de esudiar y comprender la biodiversidad de los mamíferos marinos antárticos 59 Elías Barticevic Cornejo y FOTOGRAFÍA Anelio Aguayo Lobo Óscar Barrientos Bradasic (Eds.) La desconocida vida y diversidad de los invertebrados terresres antárticos 67 Agradecemos a los autores citados en los créditos Daniel González Acuña fotográficos por su aporte en imágenes al regisro Autores Caracterísicas de la fauna que habita los fondos coseros profundos de la Antártica 75 hisórico de la ciencia antártica nacional. -

8. Primate Evolution
8. Primate Evolution Jonathan M. G. Perry, Ph.D., The Johns Hopkins University School of Medicine Stephanie L. Canington, B.A., The Johns Hopkins University School of Medicine Learning Objectives • Understand the major trends in primate evolution from the origin of primates to the origin of our own species • Learn about primate adaptations and how they characterize major primate groups • Discuss the kinds of evidence that anthropologists use to find out how extinct primates are related to each other and to living primates • Recognize how the changing geography and climate of Earth have influenced where and when primates have thrived or gone extinct The first fifty million years of primate evolution was a series of adaptive radiations leading to the diversification of the earliest lemurs, monkeys, and apes. The primate story begins in the canopy and understory of conifer-dominated forests, with our small, furtive ancestors subsisting at night, beneath the notice of day-active dinosaurs. From the archaic plesiadapiforms (archaic primates) to the earliest groups of true primates (euprimates), the origin of our own order is characterized by the struggle for new food sources and microhabitats in the arboreal setting. Climate change forced major extinctions as the northern continents became increasingly dry, cold, and seasonal and as tropical rainforests gave way to deciduous forests, woodlands, and eventually grasslands. Lemurs, lorises, and tarsiers—once diverse groups containing many species—became rare, except for lemurs in Madagascar where there were no anthropoid competitors and perhaps few predators. Meanwhile, anthropoids (monkeys and apes) emerged in the Old World, then dispersed across parts of the northern hemisphere, Africa, and ultimately South America. -

Evidence for a Convergent Slowdown in Primate Molecular Rates and Its Implications for the Timing of Early Primate Evolution
Evidence for a convergent slowdown in primate molecular rates and its implications for the timing of early primate evolution Michael E. Steipera,b,c,d,1 and Erik R. Seifferte aDepartment of Anthropology, Hunter College of the City University of New York (CUNY), New York, NY 10065; Programs in bAnthropology and cBiology, The Graduate Center, CUNY, New York, NY 10016; dNew York Consortium in Evolutionary Primatology, New York, NY; and eDepartment of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794-8081 Edited by Richard G. Klein, Stanford University, Stanford, CA, and approved February 28, 2012 (received for review November 29, 2011) A long-standing problem in primate evolution is the discord divergences—that molecular rates were exceptionally rapid in between paleontological and molecular clock estimates for the the earliest primates, and that these rates have convergently time of crown primate origins: the earliest crown primate fossils slowed over the course of primate evolution. Indeed, a conver- are ∼56 million y (Ma) old, whereas molecular estimates for the gent rate slowdown has been suggested as an explanation for the haplorhine-strepsirrhine split are often deep in the Late Creta- large differences between the molecular and fossil evidence for ceous. One explanation for this phenomenon is that crown pri- the timing of placental mammalian evolution generally (18, 19). mates existed in the Cretaceous but that their fossil remains However, this hypothesis has not been directly tested within have not yet been found. Here we provide strong evidence that a particular mammalian group. this discordance is better-explained by a convergent molecular Here we test this “convergent rate slowdown” hypothesis in rate slowdown in early primate evolution. -

Stem Members of Platyrrhini Are Distinct from Catarrhines in at Least One Derived Cranial Feature
Journal of Human Evolution 100 (2016) 16e24 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Stem members of Platyrrhini are distinct from catarrhines in at least one derived cranial feature * Ethan L. Fulwood a, , Doug M. Boyer a, Richard F. Kay a, b a Department of Evolutionary Anthropology, Duke University, Box 90383, Durham, NC 27708, USA b Division of Earth and Ocean Sciences, Nicholas School of the Environment, Duke University, Durham, NC 27708, USA article info abstract Article history: The pterion, on the lateral aspect of the cranium, is where the zygomatic, frontal, sphenoid, squamosal, Received 3 August 2015 and parietal bones approach and contact. The configuration of these bones distinguishes New and Old Accepted 2 August 2016 World anthropoids: most extant platyrrhines exhibit contact between the parietal and zygomatic bones, while all known catarrhines exhibit frontal-alisphenoid contact. However, it is thought that early stem- platyrrhines retained the apparently primitive catarrhine condition. Here we re-evaluate the condition of Keywords: key fossil taxa using mCT (micro-computed tomography) imaging. The single known specimen of New World monkeys Tremacebus and an adult cranium of Antillothrix exhibit the typical platyrrhine condition of parietal- Pterion Homunculus zygomatic contact. The same is true of one specimen of Homunculus, while a second specimen has the ‘ ’ Tremacebus catarrhine condition. When these new data are incorporated into an ancestral state reconstruction, they MicroCT support the conclusion that pterion frontal-alisphenoid contact characterized the last common ancestor of crown anthropoids and that contact between the parietal and zygomatic is a synapomorphy of Platyrrhini. -

Neotropical Primates
ISSN 1413-4703 NEOTROPICAL PRIMATES A Journal of the Neotropical Section of the IUCN/SSC Primate Specialist Group Volume 14 Number 1 January 2007 Editors Erwin Palacios Liliana Cortés-Ortiz Júlio César Bicca-Marques Eckhard Heymann Jessica Lynch Alfaro News and Books Reviews Brenda Solórzano Ernesto Rodríguez-Luna PSG Chairman Russell A. Mittermeier PSG Deputy Chairman Anthony B. Rylands SPECIES SURVIVAL COMMISSION Neotropical Primates A Journal of the Neotropical Section of the IUCN/SSC Primate Specialist Group Center for Applied Biodiversity Science Conservation International 2011 Crystal Drive, Suite 500, Arlington, VA 22202, USA ISSN 1413-4703 Abbreviation: Neotrop. Primates Editors Erwin Palacios, Conservation International – Colombia Liliana Cortés-Ortiz, Museum of Zoology, University of Michigan, Ann Arbor, MI, USA Júlio César Bicca-Marques, Pontifícia Universidad Católica do Rio Grande do Sul, Porto Alegre, Brasil Eckhard Heymann, Deutsches Primatenzentrum, Göttingen, Germany Jessica Lynch Alfaro, Washington State University, Pullman, WA, USA News and Books Reviews Brenda Solórzano, Instituto de Neuroetología, Universidad Veracruzana, Xalapa, México Ernesto Rodríguez-Luna, Instituto de Neuroetología, Universidad Veracruzana, Xalapa, México Founding Editors Anthony B. Rylands, Center for Applied Biodiversity Science, Conservation International, Arlington VA, USA Ernesto Rodríguez-Luna, Instituto de Neuroetología, Universidad Veracruzana, Xalapa, México Editorial Board Hannah M. Buchanan-Smith, University of Stirling, Stirling, Scotland, UK Adelmar F. Coimbra-Filho, Academia Brasileira de Ciências, Rio de Janeiro, Brazil Carolyn M. Crockett, Regional Primate Research Center, University of Washington, Seattle, WA, USA Stephen F. Ferrari, Universidade Federal do Sergipe, Aracajú, Brazil Russell A. Mittermeier, Conservation International, Arlington, VA, USA Marta D. Mudry, Universidad de Buenos Aires, Argentina Horácio Schneider, Universidade Federal do Pará, Campus Universitário de Bragança, Brazil Karen B. -

First North American Fossil Monkey and Early Miocene Tropical Biotic Interchange Jonathan I
LETTER doi:10.1038/nature17415 First North American fossil monkey and early Miocene tropical biotic interchange Jonathan I. Bloch1, Emily D. Woodruff1,2, Aaron R. Wood1,3, Aldo F. Rincon1,4, Arianna R. Harrington1,2,5, Gary S. Morgan6, David A. Foster4, Camilo Montes7, Carlos A. Jaramillo8, Nathan A. Jud1, Douglas S. Jones1 & Bruce J. MacFadden1 New World monkeys (platyrrhines) are a diverse part of modern Primates Linnaeus, 1758 tropical ecosystems in North and South America, yet their early Anthropoidea Mivart, 1864 evolutionary history in the tropics is largely unknown. Molecular Platyrrhini Geoffroy, 1812 divergence estimates suggest that primates arrived in tropical Cebidae Bonaparte, 1831 Central America, the southern-most extent of the North American Panamacebus transitus gen. et sp. nov. landmass, with several dispersals from South America starting with the emergence of the Isthmus of Panama 3–4 million years Etymology. Generic name combines ‘Panama’ with ‘Cebus’, root taxon ago (Ma)1. The complete absence of primate fossils from Central for Cebidae. Specific name ‘transit’ (Latin, crossing) refers to its implied America has, however, limited our understanding of their history early Miocene dispersal between South and North America. in the New World. Here we present the first description of a fossil Holotype. UF 280128, left upper first molar (M1; Fig. 2a, b). monkey recovered from the North American landmass, the oldest Referred material. Left upper second molar (M2; UF 281001; Fig. 2a, b), known crown platyrrhine, from a precisely dated 20.9-Ma layer in partial left lower first incisor (I1; UF 280130), right lower second the Las Cascadas Formation in the Panama Canal Basin, Panama. -

New Platyrrhine Tali from La Venta, Colombia Department of Anthropology, Northern Illinois University, Dekalb, Illinois 60115
Daniel L. Gebo New platyrrhine tali from La Venta, Colombia Department of Anthropology, Northern Illinois University, DeKalb, Illinois 60115. U.S.A. Two new primate tali were discovered from the middle Miocene of South America at La Venta, Colombia. IGM-KU 8802 is similar in morphology to Callicebus and Aotus, and is allocated to cf. Aotus dindensis, while IGM-KU Marian Dagosto 8803, associated with a dentition of a new cebine primate, is similar to Saimiri. Both tali differ from the other known fossil platyrrhine tali, Departments of Cell, Molecular, and Dolichocebus and Cebupithecia, and increase our knowledge of the locomotor Structural Biology and Anthropology, diversity of the La Venta primate fauna. hrorthrerestern lJniuersi;v, Euanston, Illinois 60208, U.S.A. Alfred L. Rosenberger Department of Anthropology, L’niuersity of Illinois at Chicago, Chicago, Illinois 60680, U.S.A. Takeshi Setoguchi Primate Research Institute, Kyoto C’niniuersiQ,Inuyama Cily, Aichi 484, Japan Received 18 April 1989 Revision received 4 December 1989and accepted 2 1 December 1989 Keywords: Platyrrhini, La Venta, foot bones, locomotion. Journal of Human Evolution (1990) 19,737-746 Introduction Two new platyrrhine tali were discovered at La Venta, Colombia, by the Japanese/American field team working in conjunction with INGEOMINAS (Instituto National de Investigaciones Geologico-Mineras) during the field season of 1988. These two fossils add to the rare but growing number of postcranial remains of extinct platyrrhines from the Miocene of South America (Stirton, 1951; Gebo & Simons, 1987; Anapol & Fleagle, 1988; Ford, 1990). They represent the first new primate postcranials to be described from La Venta in nearly four decades. -

La Venta Submitted to Primates 11-27-09
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Kent Academic Repository 1 Community Ecology of the Middle Miocene Primates of La Venta, 2 Colombia: the Relationship between Ecological Diversity, Divergence 3 Time, and Phylogenetic Richness 4 5 The final publication is available at Springer via http://dx.doi.org/10.1007/s10764-010-9419-1 6 7 8 Brandon C. Wheeler 9 Interdepartmental Doctoral Program in Anthropological Sciences 10 Stony Brook University 11 Stony Brook, NY 11794-4364 USA 12 Phone: 1-631-675-6412 13 Fax: 1-631-632-9165 14 E-mail: [email protected] 15 16 17 Size of the manuscript: 18 Word count (whole file): 4,622 19 Word count abstract: 229 20 3 tables & 8 figures 21 22 Originally submitted to Primates on July 15, 2009 23 Revision submitted on November 26, 2009 24 1 25 26 2 27 Abstract 28 It has been suggested that the degree of ecological diversity that characterizes a primate 29 community correlates positively with both its phylogenetic richness 30 and the time since the members of that community diverged (Fleagle and Reed 1999). It is 31 therefore questionable whether or not a community with a relatively recent divergence time 32 but high phylogenetic richness would be as ecologically variable as a community with 33 similar phylogenetic richness but a more distant divergence time. To address this question, 34 the ecological diversity of a fossil primate community from La Venta, Colombia, a Middle 35 Miocene platyrrhine community with phylogenetic diversity comparable to extant 36 platyrrhine communities but a relatively short time since divergence, was compared with 37 that of modern neotropical primate communities. -

Taxonomy, Phylogeny and Distribution of Tamarins (Genus Saguinus, Hoffmannsegg 1807)
Göttinger Zentrum für Biodiversitätsforschung und Ökologie ‐Göttingen Centre for Biodiversity and Ecology‐ TAXONOMY, PHYLOGENY AND DISTRIBUTION OF TAMARINS (GENUS SAGUINUS, HOFFMANNSEGG 1807) Dissertation zur Erlangung des Doktorgrades der Mathematisch‐Naturwissenschaftlichen Fakultäten der Georg‐August‐Universität zu Göttingen vorgelegt von Dipl.‐Biol. Christian Matauschek aus München Göttingen, Dezember 2010 Referent: Prof. Dr. Eckhard W. Heymann Koreferent: Prof. Dr. Peter M. Kappeler Tag der mündlichen Prüfung: i Contents 1 GENERAL INTRODUCTION.........................................................................................................1 1.1 THE TAMARINS OF THE GENUS SAGUINUS (HOFFMANNSEGG 1807)....................................................... 2 1.2 OVERVIEW OF THE CURRENT STATUS OF TAMARIN TAXONOMY ............................................................... 2 1.3 GEOGRAPHIC ORIGIN AND DISPERSAL OF SAGUINUS........................................................................... 10 1.4 SPECIFIC QUESTIONS .................................................................................................................... 13 2 COMPLETE MITOCHONDRIAL GENOME DATA REVEAL THE PHYLOGENY OF CALLITRICHINE PRIMATES AND A LATE MIOCENE DIVERGENCE OF TAMARIN SPECIES GROUPS ..............................15 2.1 INTRODUCTION ........................................................................................................................... 17 2.2 METHODS ................................................................................................................................. -
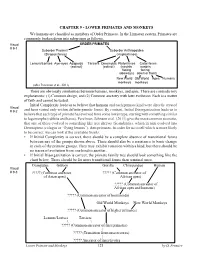
CHAPTER 9 - LOWER PRIMATES and MONKEYS We Humans Are Classified As Members of Order Primates
CHAPTER 9 - LOWER PRIMATES AND MONKEYS We humans are classified as members of Order Primates. In the Linnaean system, Primates are commonly broken down into subgroups as follows. Visual ORDER PRIMATES # 9-1 Suborder Prosimii Suborder Anthropoidea (Strepsirrhines) (Haplorrhines) Lemurs/Lorises Aye-ayes Adapoids Tarsiers Omomyids Platyrrhines Catarrhines (extinct) (extinct) (nostrils nostrils facing facing sideways) down or front) New World Old World Apes Humans monkeys monkeys (after Perelman et al., 2011) There are obviously similarities between humans, monkeys, and apes. There are contradictory explanations: (1) Common design, and (2) Common ancestry with later evolution. Each is a matter of faith and cannot be tested. Initial Complexity leads us to believe that humans and each primate kind were directly created Visual # 9-2 and have varied only within definite genetic limits. By contrast, Initial Disorganization leads us to believe that each type of primate has evolved from some lower type, starting with something similar to lagomorphs (rabbits and hares). Perelman, Johnson et al. (2011) give the most common scenario, that one of these evolved to something like tree shrews (Scandentia), which in turn evolved into Dermoptera (colugos or “flying lemurs”), then primates. In order for us to tell which is more likely to be correct, we can look at the available fossils. • If Initial Complexity is correct, there should be a complete absence of transitional forms between any of the groups shown above. There should also be a resistance to basic change in each of the primate groups. They may exhibit variation within a kind, but there should be no traces of evolution from one kind to another.