Root System Attributes of 12 Juvenile Indigenous Early Colonising Shrub and Tree Species with Potential for Mitigating Erosion in New Zealand M
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
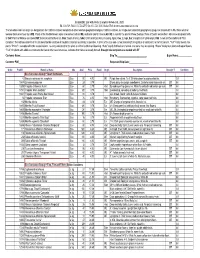
Availability List with Order Form for August 20, 2021 Ph. 800-949
Availability List with Order Form for October 01, 2021 Ph. 800-949-5064/831-728-2595/fax 831-728-3146 On the Web at www.suncrestnurseries.com The two-letter codes on origin are designations for California natives and plants of other selected geographical origins. California natives, our single most important geographical group, are marked with a CA. Those of other western states are shown by a WE. Plants of the Mediterranean region are marked with an ME. Australian plants have an AU. NZ is used for the plants of New Zealand. Those of South and southern Africa are designated with an SAf. Plants of Mexico are labeled MX; those of Central America, CAm; South America, SAm; China and East Asia, including Japan, Cea; Europe, Eur; and plants of hybrid origin, Hbd. A note on Descriptions of Plant Condition: The last two columns of this list describe the condition of the plants listed as accurately as possible, in terms of the size and current blooming/fruiting status of each plant. In the first column, "Full" = fully rooted, nice plants. "Prime" = exceptional! In the second column, no entry indicates that the plant is neither budded nor blooming. “Buds” signify that buds are formed, and some may be opening. “Bloom” designates plants with open flowers. “Fruit”, for plants with edible or ornamental berries or other seed structures, indicates that fruits are already formed. Drought tolerant plants are marked with DT Customer Name:______________________________ Ship To:_____________________________ Buyer Name:_____________ Customer PO#______________________________ Requested Ship Date:____________________ Order ProdID Botanical Name Size Avail Price Retail Origin Description Drought T Condition Eco Conscious Beauty™/Quart Containers 43 Brassica oleracea var. -

<I>Plagianthus</I> (Malveae, Malvaceae)
Systematic Botany (2011), 36(2): pp. 405–418 © Copyright 2011 by the American Society of Plant Taxonomists DOI 10.1600/036364411X569589 Phylogeny and Character Evolution in the New Zealand Endemic Genus Plagianthus (Malveae, Malvaceae) Steven J. Wagstaff 1 , 3 and Jennifer A. Tate 2 1 Allan Herbarium, Landcare Research, PO Box 40, Lincoln 7640, New Zealand 2 Massey University, Institute of Molecular Biosciences, Private Bag 11222, Palmerston North, New Zealand 3 Author for correspondence ([email protected]) Communicating Editor: Lúcia Lohmann Abstract— As presently circumscribed, Plagianthus includes two morphologically distinct species that are endemic to New Zealand. Plagianthus divaricatus , a divaricate shrub, is a dominant species in coastal saline shrub communities, whereas P. regius is a tree of lowland and montane forests. Results from independent analyses of ITS and 5′ trnK / matK sequences are congruent, and when combined provide a robust framework to study character evolution. Our findings suggest the ancestor of Plagianthus originated in Australia where the sister gen- era Asterotrichion and Gynatrix are presently distributed. The stem age of Plagianthus was estimated at 7.3 (4.0–14.0) million years ago (Ma) and the crown radiation at 3.9 (1.9–8.2) Ma. Most of the characters optimized onto the molecular phylogeny were shared with source lineages from Australia and shown to be plesiomorphic. Only the divaricate branching pattern characteristic of Plagianthus divaricatus was acquired after the lineage became established in New Zealand and shown to be apomorphic. The initial Plagianthus founders were shrubs or small trees with deciduous leaves and small inconspicuous dioecious flowers. -

Bio 308-Course Guide
COURSE GUIDE BIO 308 BIOGEOGRAPHY Course Team Dr. Kelechi L. Njoku (Course Developer/Writer) Professor A. Adebanjo (Programme Leader)- NOUN Abiodun E. Adams (Course Coordinator)-NOUN NATIONAL OPEN UNIVERSITY OF NIGERIA BIO 308 COURSE GUIDE National Open University of Nigeria Headquarters 14/16 Ahmadu Bello Way Victoria Island Lagos Abuja Office No. 5 Dar es Salaam Street Off Aminu Kano Crescent Wuse II, Abuja e-mail: [email protected] URL: www.nou.edu.ng Published by National Open University of Nigeria Printed 2013 ISBN: 978-058-434-X All Rights Reserved Printed by: ii BIO 308 COURSE GUIDE CONTENTS PAGE Introduction ……………………………………......................... iv What you will Learn from this Course …………………............ iv Course Aims ……………………………………………............ iv Course Objectives …………………………………………....... iv Working through this Course …………………………….......... v Course Materials ………………………………………….......... v Study Units ………………………………………………......... v Textbooks and References ………………………………........... vi Assessment ……………………………………………….......... vi End of Course Examination and Grading..................................... vi Course Marking Scheme................................................................ vii Presentation Schedule.................................................................... vii Tutor-Marked Assignment ……………………………….......... vii Tutors and Tutorials....................................................................... viii iii BIO 308 COURSE GUIDE INTRODUCTION BIO 308: Biogeography is a one-semester, 2 credit- hour course in Biology. It is a 300 level, second semester undergraduate course offered to students admitted in the School of Science and Technology, School of Education who are offering Biology or related programmes. The course guide tells you briefly what the course is all about, what course materials you will be using and how you can work your way through these materials. It gives you some guidance on your Tutor- Marked Assignments. There are Self-Assessment Exercises within the body of a unit and/or at the end of each unit. -

A Landscape-Based Assessment of Climate Change Vulnerability for All Native Hawaiian Plants
Technical Report HCSU-044 A LANDscape-bASED ASSESSMENT OF CLIMatE CHANGE VULNEraBILITY FOR ALL NatIVE HAWAIIAN PLANts Lucas Fortini1,2, Jonathan Price3, James Jacobi2, Adam Vorsino4, Jeff Burgett1,4, Kevin Brinck5, Fred Amidon4, Steve Miller4, Sam `Ohukani`ohi`a Gon III6, Gregory Koob7, and Eben Paxton2 1 Pacific Islands Climate Change Cooperative, Honolulu, HI 96813 2 U.S. Geological Survey, Pacific Island Ecosystems Research Center, Hawaii National Park, HI 96718 3 Department of Geography & Environmental Studies, University of Hawai‘i at Hilo, Hilo, HI 96720 4 U.S. Fish & Wildlife Service —Ecological Services, Division of Climate Change and Strategic Habitat Management, Honolulu, HI 96850 5 Hawai‘i Cooperative Studies Unit, Pacific Island Ecosystems Research Center, Hawai‘i National Park, HI 96718 6 The Nature Conservancy, Hawai‘i Chapter, Honolulu, HI 96817 7 USDA Natural Resources Conservation Service, Hawaii/Pacific Islands Area State Office, Honolulu, HI 96850 Hawai‘i Cooperative Studies Unit University of Hawai‘i at Hilo 200 W. Kawili St. Hilo, HI 96720 (808) 933-0706 November 2013 This product was prepared under Cooperative Agreement CAG09AC00070 for the Pacific Island Ecosystems Research Center of the U.S. Geological Survey. Technical Report HCSU-044 A LANDSCAPE-BASED ASSESSMENT OF CLIMATE CHANGE VULNERABILITY FOR ALL NATIVE HAWAIIAN PLANTS LUCAS FORTINI1,2, JONATHAN PRICE3, JAMES JACOBI2, ADAM VORSINO4, JEFF BURGETT1,4, KEVIN BRINCK5, FRED AMIDON4, STEVE MILLER4, SAM ʽOHUKANIʽOHIʽA GON III 6, GREGORY KOOB7, AND EBEN PAXTON2 1 Pacific Islands Climate Change Cooperative, Honolulu, HI 96813 2 U.S. Geological Survey, Pacific Island Ecosystems Research Center, Hawaiʽi National Park, HI 96718 3 Department of Geography & Environmental Studies, University of Hawaiʽi at Hilo, Hilo, HI 96720 4 U. -

WRA Species Report
Family: Malvaceae Taxon: Lagunaria patersonia Synonym: Hibiscus patersonius Andrews Common Name: cowitchtree Lagunaria patersonia var. bracteata Benth. Norfolk Island-hibiscus Lagunaria queenslandica Craven Norfolk-hibiscus pyramid-tree sallywood white-oak whitewood Questionaire : current 20090513 Assessor: Patti Clifford Designation: H(HPWRA) Status: Assessor Approved Data Entry Person: Patti Clifford WRA Score 7 101 Is the species highly domesticated? y=-3, n=0 n 102 Has the species become naturalized where grown? y=1, n=-1 103 Does the species have weedy races? y=1, n=-1 201 Species suited to tropical or subtropical climate(s) - If island is primarily wet habitat, then (0-low; 1-intermediate; 2- High substitute "wet tropical" for "tropical or subtropical" high) (See Appendix 2) 202 Quality of climate match data (0-low; 1-intermediate; 2- High high) (See Appendix 2) 203 Broad climate suitability (environmental versatility) y=1, n=0 y 204 Native or naturalized in regions with tropical or subtropical climates y=1, n=0 y 205 Does the species have a history of repeated introductions outside its natural range? y=-2, ?=-1, n=0 y 301 Naturalized beyond native range y = 1*multiplier (see y Appendix 2), n= question 205 302 Garden/amenity/disturbance weed n=0, y = 1*multiplier (see Appendix 2) 303 Agricultural/forestry/horticultural weed n=0, y = 2*multiplier (see n Appendix 2) 304 Environmental weed n=0, y = 2*multiplier (see y Appendix 2) 305 Congeneric weed n=0, y = 1*multiplier (see n Appendix 2) 401 Produces spines, thorns or burrs y=1, n=0 -

An Additional Nomenclatural Transfer in the Pantropical Genus Myrsine (Primulaceae: Myrsinoideae) John J
Nova Southeastern University NSUWorks Marine & Environmental Sciences Faculty Articles Department of Marine and Environmental Sciences 9-13-2018 An Additional Nomenclatural Transfer in the Pantropical Genus Myrsine (Primulaceae: Myrsinoideae) John J. Pipoly III Broward County Parks & Recreation Division; Nova Southeastern University, [email protected] Jon M. Ricketson Missouri Botanical Garden Find out more information about Nova Southeastern University and the Halmos College of Natural Sciences and Oceanography. Follow this and additional works at: https://nsuworks.nova.edu/occ_facarticles Part of the Marine Biology Commons, and the Oceanography and Atmospheric Sciences and Meteorology Commons NSUWorks Citation John J. Pipoly III and Jon M. Ricketson. 2018. An Additional Nomenclatural Transfer in the Pantropical Genus Myrsine (Primulaceae: Myrsinoideae) .Novon , (3) : 287 -287. https://nsuworks.nova.edu/occ_facarticles/942. This Article is brought to you for free and open access by the Department of Marine and Environmental Sciences at NSUWorks. It has been accepted for inclusion in Marine & Environmental Sciences Faculty Articles by an authorized administrator of NSUWorks. For more information, please contact [email protected]. An Additional Nomenclatural Transfer in the Pantropical Genus Myrsine (Primulaceae: Myrsinoideae) John J. Pipoly III Broward County Parks & Recreation Division, 950 NW 38th St., Oakland Park, Florida 33309, U.S.A.; Nova Southeastern University, 8000 N Ocean Dr., Dania Beach, Florida 33004, U.S.A. [email protected]; [email protected]; [email protected] Jon M. Ricketson Missouri Botanical Garden, 4344 Shaw Blvd., St. Louis, Missouri 63110, U.S.A. [email protected] ABSTRACT. Rapanea pellucidostriata Gilg & Schellenb. TYPE: Democratic Republic of the Congo. Ruwen- is transferred to Myrsine L. -

Phytochemical Analysis of Fruit Extract of Myrsine Africana
International Journal of Pharmacy and Pharmaceutical Sciences Academic Sciences ISSN- 0975-1491 Vol 3, Issue 4, 2011 Research Article PHYTOCHEMICAL ANALYSIS OF FRUIT EXTRACT OF MYRSINE AFRICANA VASUDHA ABBHI*, LINCY JOSEPH, MATHEW GEORGE School of Pharmaceutical Sciences, Shoolini University, Solan (H.P.) India. Email: [email protected] Received: 13 Aug 2011, Revised and Accepted: 11 Sep 2011 ABSTRACT The present paper deals with the phytochemical screening of therapeutic importance from Myrsine africana, an important medicinal plant. This study involves the preliminary screening and quantitative determination of secondary metabolites from the fruits of M.africana. The phytochemical analysis revealed the presence of saponins, tannins, flavonoids, amino acids, steroids and reducing sugar. The amount of the saponins and tannins in methanolic extracts are reported (17.5% and 4% respectively). The generated data has provided the basis for its wide use as the therapeutant both in the traditional and folk medicines. Keywords: Myrsine Africana, Fruits, Phytochemical analysis. INTRODUCTION Horticulture and Forestry, Nauni, Solan (H.P.). Voucher specimens were deposited with the Herbarium at Nauni and are entered in the In recent times, there have been increased waves of interest in the UHF-Herbarium Field book no. 5585 dated 14.09.2010. field of Research in Natural Products Chemistry1. Plants have been used as treatments for thousands of years, based on experience and Preparation of extracts folk remedies and continue to draw wide attention for their role in the treatment of mild and chronic diseases2. The plant kingdom Aqueous extract represents an enormous reservoir of biologically active compounds The aqueous extract of fruits of Myrsine africana was prepared by with various chemical structures and protective/disease preventive the method of Decoction: properties (phytochemicals). -

From New Guinea
Phytotaxa 442 (3): 133–137 ISSN 1179-3155 (print edition) https://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2020 Magnolia Press Article ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.442.3.1 A new species of Myrsine (Primulaceae-Myrsinoideae) from New Guinea TIMOTHY M.A. UTTERIDGE1,3 & BRENDAN J. LEPSCHI2,4 1 Identification & Naming Dept, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AE, UK. 2 Australian National Herbarium, Centre for Australian National Biodiversity Research, GPO Box 1700, Canberra, ACT, 2601, Australia. 3 [email protected]; https://orcid.org/0000-0003-2823-0337 4 [email protected]; https://orcid.org/0000-0002-3281-2973 Abstract Myrsine exquisitorum Utteridge & Lepschi (Primulaceae-Myrsinoideae) is described and illustrated as a new species endemic to the Western Highlands Province from Papua New Guinea. The new species is unique in the relatively large, almost orbicular leaves with entire margins, and the tetramerous flowers arranged in axillary fascicles without forming short shoots. Keywords: Papuasia, Malesia, Myrsinaceae, Rapanea, taxonomy Introduction Myrsine Linnaeus (1753: 196) is a pantropical genus of c. 280 species, and now includes all taxa previously placed in the paleotropical genus Rapanea Aublet (1775: 121) and the New World genus Suttonia A.Richard (1832: 349). Traditionally, Rapanea was considered distinct from Myrsine based on the absence of well-defined filaments and style, but these characters have been shown to intergrade between species of the different genera. Pipoly (2007) and Takeuchi & Pipoly (2009) further discuss character variation in the genera in New Guinea, and formally completed the transfer of the eastern Malesian names of Rapanea to Myrsine. -

Familia Pittosporaceae
© 2012. José Manuel Sánchez de Lorenzo‐Cáceres FAMILIA PITTOSPORACEAE Árboles, arbustos siempre verdes, a veces con tallos las anteras ditecas, introrsas, que abren por suturas volubles o espinosos, con o sin indumento. Hojas simples, longitudinales o más raramente por poros apicales. Disco alternas o pseudoverticiladas hacia el extremo de los nectarífero ausente. Gineceo sincárpico, ausente en las tallos, generalmente enteras, de textura gruesa, sin flores masculinas, formado por 2 (‐5) carpelos unidos estípulas. Inflorescencias axilares o terminales, cimosas, formando un ovario súpero, sésil o cortamente estipitado, corimbosas o flores solitarias, con brácteas y bractéolas. unilocular, más raramente plurilocular, con varios a Flores bisexuales, a veces funcionalmente unisexuales, numerosos rudimentos seminales en cada lóculo. Estilo generalmente actinomorfas, hipóginas. Cáliz con 5 simple y corto, terminal, con estigma pequeño, capitado o sépalos, libres o soldados basalmente, imbricados, lobulado. Fruto en cápsula loculicida o en baya, con regulares; corola con 5 pétalos, libres o ligeramente numerosas semillas inmersas a menudo en una pulpa unidos en la base, imbricados, unguiculados. Androceo de pegajosa. Comprende 9 géneros y alrededor de 240 5 estambres fértiles, ausentes en las flores femeninas, especies distribuidas por los trópicos y regiones templadas alternando con los pétalos, libres o unidos en la base, con del Viejo Mundo. Pittosporum tobira Pittosporum tobira ‘Variegatum’ Pittosporum angustifolium Pittosporum truncatum Pittosporum truncatum ‘Variegatum’ Pittosporum tenuifolium Pittosporum tenuifolium Gaertn. = Pittosporum mayi hort. Arbusto o pequeño arbolito con los años, de hojas a menudo con los bordes algo ondulados y flores purpúreas, nativo de Nueva Zelanda. Además de la forma típica, a veces se Plantas ornamentales de los jardines de Murcia. -

VH Flora Complete Rev 18-19
Flora of Vinalhaven Island, Maine Macrolichens, Liverworts, Mosses and Vascular Plants Javier Peñalosa Version 1.4 Spring 2019 1. General introduction ------------------------------------------------------------------------1.1 2. The Setting: Landscape, Geology, Soils and Climate ----------------------------------2.1 3. Vegetation of Vinalhaven Vegetation: classification or description? --------------------------------------------------3.1 The trees and shrubs --------------------------------------------------------------------------3.1 The Forest --------------------------------------------------------------------------------------3.3 Upland spruce-fir forest -----------------------------------------------------------------3.3 Deciduous woodlands -------------------------------------------------------------------3.6 Pitch pine woodland ---------------------------------------------------------------------3.6 The shore ---------------------------------------------------------------------------------------3.7 Rocky headlands and beaches ----------------------------------------------------------3.7 Salt marshes -------------------------------------------------------------------------------3.8 Shrub-dominated shoreline communities --------------------------------------------3.10 Freshwater wetlands -------------------------------------------------------------------------3.11 Streams -----------------------------------------------------------------------------------3.11 Ponds -------------------------------------------------------------------------------------3.11 -

An Evaluation of the Establishment, Early Growth, and Nutritive Value of Native New Zealand Shrubs'
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the permission of the Author. An evaluation of the establishment, early growth, and nutritive value of native New Zealand shrubs' A thesis presented in partial fulfilment of the requirements for the degree of Masters In Environmental Management At Massey University, Palmerston North, New Zealand Georgia Rose Simmonds 2020 Abstract Agriculture in New Zealand faces many challenges including a need to develop more environmentally focused production systems to help address issues including the need to revegetate step erosion prone hill country, improve indigenous biodiversity and improve water quality. In the past New Zealand has experienced increasingly unpredictable and severe weather which has resulted in severe damage, for example, the 2004 flooding event in the lower north island (Fuller, 2005). New Zealand has a large portion of land that is classed as hill country or steep land, much of which is also classed as highly erodible. These highly erodible areas are vulnerable to high intensity rainfall events; revegetation could help mitigate or reduce the effects of erosion. The species that are currently used in erosion control on farms are often poplars and willows due to their ability to stabilise hill slopes and ease of planting. Native species are not often used in erosion control projects, potentially due to planting difficulties but more likely due to a lack of consistent and long-term information (Phillips, 2005). -

Here Before We Humans Were and Their Relatives Will Probably Be Here When We Are Gone
The ‘mighty tōtara’ is one of our most extraordinary trees. Among the biggest and oldest trees in the New Zealand forest, the heart of Māori carving and culture, trailing no. 8 wire as fence posts on settler farms, clambered up in the Pureora protests of the 1980s: the story of New Zealand can be told through tōtara. Simpson tells that story like nobody else could. In words and pictures, through waka and leaves, farmers and carvers, he takes us deep inside the trees: their botany and evolution, their role in Māori life and lore, their uses by Pākehā, and their current status in our environment and culture. By doing so, Simpson illuminates the natural world and the story of Māori and Pākehā in this country. Our largest trees, the kauri Tāne Mahuta and the tōtara Pouakani, are both thought to be around 1000 years old. They were here before we humans were and their relatives will probably be here when we are gone. Tōtara has been central to life in this country for thousands of years. This book tells a great tree’s story, and that is our story too. Philip Simpson is a botanist and author of Dancing Leaves: The Story of New Zealand’s Cabbage Tree, Tī Kōuka (Canterbury University Press, 2000) and Pōhutukawa and Rātā: New Zealand’s Iron-hearted Trees (Te Papa Press, 2005). Both books won Montana Book Awards in the Environment category and Pōhutukawa and Rātā also won the Montana Medal for best non-fiction book. Simpson is unique in his ability to combine the scientific expertise of the trained botanist with a writer’s ability to understand the history of Māori and Pākehā interactions with the environment.