Conditioned Behavior in Drosophila Melanogaster (Learnin/Memory/Odor Discrimination/Color Vision)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
NIHAA Summer 1993
The Newsletter of the NIH Alumni Association Summer 1993 Vol. 5, No . 2 date Nobel Laureate Harold Varmus Nominated as 14th NIH Director Ruth Kirschstein Named Acting Director President Clinton on A ug. 3 announced his intention to nominate Dr. Harold Eliot Varmus as the 14th director of th e National Institutes of Health. A Senate confirmation process must precede Yarmus· taking over leadership of the institutes. Winner of the Nobel Prize in 1989 for his work in cancer research. Va1111us. 53. is a professor of microbi ology. biochemistry. and biophysics. and the American Cancer Society pro fessor vfmoh:cu/ur >1iro/O£)' iJI l/Je Uni versity of California, San Francisco. He is a leader in the stu dy of cancer causing genes called "oncogenes," and an intemationall-y fecogni:z.ed authof\t-y Dr. Ruth l. Kirschsteln , acting NIH director on retroviruses. the viruses that cause Dr. Harold E. Varmus , direclor-designale AIDS and many cancers in animnl.. FIC 25 Years Old In '93 Thirty-eight-year NIH veteran Dr. Research Festival '93 Schedule Ruth Kirschstein. director of NIGM S Scholars-in-Residence (See Director p. 6) NIHAA Members Invited Program Celebrates To Alumni Symposium In This Issue Tile fas\ morning ofNCH Rc:-.c;\rch Nursing cell/er /J1·1·111111•s Festival '93-Monday. Sept. 20-has This year. the Fogarty Intern ational 17th i11s1it11tc• 111 NII/ p. ? been designated National lnsritute of Center (FIC) is 25 years old . T he cen Greeri11gs from 1/,11 1w11• NII /AA prl'sidt'lll. Diabetes and Digestive and Kidney Tltolll(IS .I. -

Advertising (PDF)
Neuroscience 2013 SEE YOU IN San Diego November 9 – 13, 2013 Join the Society for Neuroscience Are you an SfN member? Join now and save on annual meeting registration. You’ll also enjoy these member-only benefits: • Abstract submission — only SfN members can submit abstracts for the annual meeting • Lower registration rates and more housing choices for the annual meeting • The Journal of Neuroscience — access The Journal online and receive a discounted subscription on the print version • Free essential color charges for The Journal of Neuroscience manuscripts, when first and last authors are members • Free online access to the European Journal of Neuroscience • Premium services on NeuroJobs, SfN’s online career resource • Member newsletters, including Neuroscience Quarterly and Nexus If you are not a member or let your membership lapse, there’s never been a better time to join or renew. Visit www.sfn.org/joinnow and start receiving your member benefits today. www.sfn.org/joinnow membership_full_page_ad.indd 1 1/25/10 2:27:58 PM The #1 Cited Journal in Neuroscience* Read The Journal of Neuroscience every week to keep up on what’s happening in the field. s4HENUMBERONECITEDJOURNAL INNEUROSCIENCE s4HEMOSTNEUROSCIENCEARTICLES PUBLISHEDEACHYEARNEARLY in 2011 s )MPACTFACTOR s 0UBLISHEDTIMESAYEAR ,EARNMOREABOUTMEMBERAND INSTITUTIONALSUBSCRIPTIONSAT *.EUROSCIORGSUBSCRIPTIONS *ISI Journal Citation Reports, 2011 The Journal of Neuroscience 4HE/FlCIAL*OURNALOFTHE3OCIETYFOR.EUROSCIENCE THE HISTORY OF NEUROSCIENCE IN AUTOBIOGRAPHY THE LIVES AND DISCOVERIES OF EMINENT SENIOR NEUROSCIENTISTS CAPTURED IN AUTOBIOGRAPHICAL BOOKS AND VIDEOS The History of Neuroscience in Autobiography Series Edited by Larry R. Squire Outstanding neuroscientists tell the stories of their scientific work in this fascinating series of autobiographical essays. -

Cold Spring Harbor Symposia on Quantitative Biology
COLD SPRING HARBOR SYMPOSIA ON QUANTITATIVE BIOLOGY VOLUME XL COLD SPRING HARBOR SYMPOSIA 0 N Q UA NTITA T1VE BIOLO G Y Founded in 1933 by REGINALD G. HARRIS Director of the Biological Laboratory 1924 to 1936 Volume I (1933).Surface Phenomena Volume II (1934) Aspects of Growth Volume III (1935) Photochemical Reactions Volume IV (1936) Excitation Phenomena Volume V (1937) Internal Secretions Volume VI (1938) Protein Chemistry Volume VII (1939) Biological Oxidations Volume VIII (1940) Permeability and the Nature of Cell Membranes Volume IX (1941) Genes and Chromosomes: Structure and Organization Volume X (1942) The Relation of Hormones to Development Volume XI (1946) Heredity and Variation in Microorganisms Volume XII (1947) Nucleic Acids and Nucleoproteins Volume XIII (1948) Biological Applications of Tracer Elements Volume XIV (1949) Amino Acids and Proteins Volume XV (1950) Origin and Evolution of Man Volume XVI (1951) Genes and Mutations Volume XVII (1952) The Neuron Volume XVIII (1953) Viruses Volume XIX (1954) The Mammalian Fetus: Physiological Aspects of Development Volume XX (1955) Population Genetics: The Nature and Causes of Genetic Variability in Population Volume XXI (1956) Genetic Mechanisms: Structure and Function Volume XXII (1957) Population Studies: Animal Ecology and Demography Volume XXIII (1958) Exchange of Genetic Material: Mechanism and Consequences Volume XXIV (1959) Genetics and Twentieth Century Darwinism Volume XXV (1960) Biological Clocks Volume XXVI (1961) Cellular Regulatory Mechanisms Volume XXVII (1962) Basic -

Introduction and Historical Perspective
Chapter 1 Introduction and Historical Perspective “ Nothing in biology makes sense except in the light of evolution. ” modified by the developmental history of the organism, Theodosius Dobzhansky its physiology – from cellular to systems levels – and by the social and physical environment. Finally, behaviors are shaped through evolutionary forces of natural selection OVERVIEW that optimize survival and reproduction ( Figure 1.1 ). Truly, the study of behavior provides us with a window through Behavioral genetics aims to understand the genetic which we can view much of biology. mechanisms that enable the nervous system to direct Understanding behaviors requires a multidisciplinary appropriate interactions between organisms and their perspective, with regulation of gene expression at its core. social and physical environments. Early scientific The emerging field of behavioral genetics is still taking explorations of animal behavior defined the fields shape and its boundaries are still being defined. Behavioral of experimental psychology and classical ethology. genetics has evolved through the merger of experimental Behavioral genetics has emerged as an interdisciplin- psychology and classical ethology with evolutionary biol- ary science at the interface of experimental psychology, ogy and genetics, and also incorporates aspects of neuro- classical ethology, genetics, and neuroscience. This science ( Figure 1.2 ). To gain a perspective on the current chapter provides a brief overview of the emergence of definition of this field, it is helpful -

2004 Albert Lasker Nomination Form
albert and mary lasker foundation 110 East 42nd Street Suite 1300 New York, ny 10017 November 3, 2003 tel 212 286-0222 fax 212 286-0924 Greetings: www.laskerfoundation.org james w. fordyce On behalf of the Albert and Mary Lasker Foundation, I invite you to submit a nomination Chairman neen hunt, ed.d. for the 2004 Albert Lasker Medical Research Awards. President mrs. anne b. fordyce The Awards will be offered in three categories: Basic Medical Research, Clinical Medical Vice President Research, and Special Achievement in Medical Science. This is the 59th year of these christopher w. brody Treasurer awards. Since the program was first established in 1944, 68 Lasker Laureates have later w. michael brown Secretary won Nobel Prizes. Additional information on previous Lasker Laureates can be found jordan u. gutterman, m.d. online at our web site http://www.laskerfoundation.org. Representative Albert Lasker Medical Research Awards Program Nominations that have been made in previous years may be updated and resubmitted in purnell w. choppin, m.d. accordance with the instructions on page 2 of this nomination booklet. daniel e. koshland, jr., ph.d. mrs. william mccormick blair, jr. the honorable mark o. hatfied Nominations should be received by the Foundation no later than February 2, 2004. Directors Emeritus A distinguished panel of jurors will select the scientists to be honored. The 2004 Albert Lasker Medical Research Awards will be presented at a luncheon ceremony given by the Foundation in New York City on Friday, October 1, 2004. Sincerely, Joseph L. Goldstein, M.D. Chairman, Awards Jury Albert Lasker Medical Research Awards ALBERT LASKER MEDICAL2004 RESEARCH AWARDS PURPOSE AND DESCRIPTION OF THE AWARDS The major purpose of these Awards is to recognize and honor individuals who have made signifi- cant contributions in basic or clinical research in diseases that are the main cause of death and disability. -

Transforming Lives
Brooklyn College Foundation Annual Report 2007–2008 Transforming Lives The First Step The doors to Brooklyn College are the doors to opportunity. Compared with other institutions of higher education, a great many of our students shoulder substantial responsibilities, and many are the first in their family to attend college. Diverse in background, interests, and ambition, they share the certainty that higher education is the way to a productive and rewarding future. For many, that future will be secured with the help of the Brooklyn College Foundation Dear Friends of Brooklyn College For students—past and present—Brooklyn College stands as a gateway to a rewarding life. They come because they want to become effective leaders in their chosen profession and engaged citizens of the world. They come because they have heard of our commitment to academic quality and to helping them reach their goals. This commitment is at the heart of who we are and what we do. We have held firm to this principle throughout my presidency and, as I leave Brooklyn College this summer, I am especially proud of what we have done together to give it life and to sustain it. Last fall, we admitted a freshman class larger and better than the year before and we were joined by forty new faculty members, bringing the number of scholars and artists we have recruited over the last nine years to 273, more than half the teaching faculty and more than we appointed in the previous three decades. We also welcomed a new Provost, Dr. William A. Tramontano, who brings proven leadership in initiating and implementing new academic programs. -

Advertising (PDF)
Edited by John Cairns, Cold Spring Harbor Laboratory; Gunther S. Stent, University of California, Berkeley; James D. Watson, Harvard University his landmark collection of essays, written by pioneers in the field of molecular biology, was first published T in 1966 as a 60th birthday tribute to Max Delbrück, a formative influence on many of today’s leading sci- entists. The book served as a valuable historical record as well as a remarkable portrait of a small, pioneering, close- knit scientific community that focused intensely on the most important questions in biology. This centennial edi- tion reflects a more personal side of Delbrück, and includes a series of photographs from his family’s albums. “Delbrück had been a kind of Gandhi of biology...It was his extraordinary personality that made him the spiritu- al force which affected the scientific and personal lives of so many people.” —Gunther S. Stent 2007, 394 pp., illus., timeline, photo gallery Hardcover $29 ISBN 978-087969800-3 CONTENTS Note from the Publisher The Injection of DNA into Cells Bacterial Conjugation Quantitatitve Tumor Virology Preface to the First Edition by Phage Elie L. Wollman H. Rubin John Cairns, Gunther S. Stent, James A.D. Hershey Story and Structure of the The Natural Selection Theory D. Watson Transfer of Parental Material to λ Transducing Phage of Antibody Formation; Ten Preface to the Expanded Edition Progeny J. Weigle Years Later John Cairns Lloyd M. Kozloff V. DNA Niels K. Jerne Electron Microscopy of Developing Growing Up in the Phage Group Cybernetics of the Insect I. ORIGINS OF MOLECULAR Optomotor Response BIOLOGY Bacteriophage J.D. -

From Controlling Elements to Transposons: Barbara Mcclintock and the Nobel Prize Nathaniel C
454 Forum TRENDS in Biochemical Sciences Vol.26 No.7 July 2001 Historical Perspective From controlling elements to transposons: Barbara McClintock and the Nobel Prize Nathaniel C. Comfort Why did it take so long for Barbara correspondence. From these and other to prevent her controlling elements from McClintock (Fig. 1) to win the Nobel Prize? materials, we can reconstruct the events moving because their effects were difficult In the mid-1940s, McClintock discovered leading up to the 1983 prize*. to study when they jumped around. She genetic transposition in maize. She What today are known as transposable never had any inclination to pursue the published her results over several years elements, McClintock called ‘controlling biochemistry of transposition. and, in 1951, gave a famous presentation elements’. During the years 1945–1946, at Current understanding of how gene at the Cold Spring Harbor Symposium, the Carnegie Dept of Genetics, Cold activity is regulated, of course, springs yet it took until 1983 for her to win a Nobel Spring Harbor, McClintock discovered a from the operon, François Jacob and Prize. The delay is widely attributed to a pair of genetic loci in maize that seemed to Jacques Monod’s 1960 model of a block of combination of gender bias and gendered trigger spontaneous and reversible structural genes under the control of an science. McClintock’s results were not mutations in what had been ordinary, adjacent set of regulatory genes (Fig. 2). accepted, the story goes, because women stable alleles. In the term of the day, they Though subsequent studies revealed in science are marginalized, because the made stable alleles into ‘mutable’ ones. -

Lasker Interactive Research Nom'18.Indd
THE 2018 LASKER MEDICAL RESEARCH AWARDS Nomination Packet albert and mary lasker foundation November 1, 2017 Greetings: On behalf of the Albert and Mary Lasker Foundation, I invite you to submit a nomination for the 2018 Lasker Medical Research Awards. Since 1945, the Lasker Awards have recognized the contributions of scientists, physicians, and public citizens who have made major advances in the understanding, diagnosis, treatment, cure, and prevention of disease. The Medical Research Awards will be offered in three categories in 2018: Basic Research, Clinical Research, and Special Achievement. The Lasker Foundation seeks nominations of outstanding scientists; nominations of women and minorities are encouraged. Nominations that have been made in previous years are not automatically reconsidered. Please see the Nomination Requirements section of this booklet for instructions on updating and resubmitting a nomination. The Foundation accepts electronic submissions. For information on submitting an electronic nomination, please visit www.laskerfoundation.org. Lasker Awards often presage future recognition of the Nobel committee, and they have become known popularly as “America’s Nobels.” Eighty-seven Lasker laureates have received the Nobel Prize, including 40 in the last three decades. Additional information on the Awards Program and on Lasker laureates can be found on our website, www.laskerfoundation.org. A distinguished panel of jurors will select the scientists to be honored with Lasker Medical Research Awards. The 2018 Awards will -
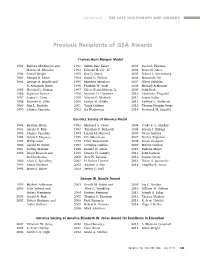
Previous Recipients of GSA Awards
GENETICS | THE 2015 GSA HONORS AND AWARDS Previous Recipients of GSA Awards Thomas Hunt Morgan Medal 1981 Barbara McClintock and 1991 Armin Dale Kaiser 2003 David S. Hogness Marcus M. Rhoades 1992 Edward H. Coe, Jr. 2004 Bruce N. Ames 1982 Sewall Wright 1993 Ray D. Owen 2005 Robert L. Metzenberg 1983 Edward B. Lewis 1994 David D. Perkins 2006 Masatoshi Nei 1984 George W. Beadle and 1995 Matthew Meselson 2007 Oliver Smithies R. Alexander Brink 1996 Franklin W. Stahl 2008 Michael Ashburner 1985 Herschel L. Roman 1997 Oliver Evans Nelson, Jr. 2009 John Roth 1986 Seymour Benzer 1998 Norman H. Horowitz 2010 Alexander Tzagoloff 1987 James F. Crow 1999 Salome G. Waelsch 2011 James Haber 1988 Norman H. Giles 2000 Evelyn M. Witkin 2012 Kathryn V. Anderson 1989 Dan L. Lindsley 2001 Yasuji Oshima 2013 Thomas Douglas Petes 1990 Charles Yanofsky 2002 Ira Herskowitz 2014 Frederick M. Ausubel Genetics Society of America Medal 1981 Beatrice Mintz 1992 Maynard V. Olson 2004 Trudy F. C. Mackay 1982 Gerald R. Fink 1993 Jonathan R. Beckwith 2005 Steven J. Elledge 1983 Charles Yanofsky 1994 Leland H. Hartwell 2006 Victor Ambros 1984 David S. Hogness 1995 Eric Wieschaus 2007 Shirley Tilghman 1985 Philip Leder 1996 Elliot Meyerowitz 2008 Susan Lindquist 1986 Gerald M. Rubin 1997 Christine Guthrie 2009 Marian Carlson 1987 Sydney Brenner 1998 Ronald W. Davis 2010 Barbara Meyer 1988 David Botstein and 1999 Charles H. Langley 2011 John Carlson Ira Herskowitz 2000 Jack W. Szostak 2012 Joanne Chory 1989 Allan C. Spradling 2001 H. Robert Horvitz 2013 Elaine A. Ostrander 1990 Nancy Kleckner 2002 Andrew Z. -

Dna Learning Center
DNA LEARNING CENTER DNA LEARNING CENTER ADMINISTRATION INSTRUCTION MULTIMEDIA David Micklos Scott Bronson Susan Lauter Judy Cumella-Korabik Amanda McBrien Shirley Chan Nancy Daidola Danielle Sixsmith Chun-hua Yang Vin Torti Veronique Bourdeau Susan Conova Elna Carrasco Uwe Hilgert Maureen Cowan We stand at the threshold of a new century with the whole human genome stretched out before us. Messages from science and the popular media suggest a world of seemingly limitless opportunities to improve human health and productivity. Yet, at the turn of the last century, science and society faced a similar rush to exploit human genetics. The story of eugenics—humankind’s first venture into a “gene age”—holds a cautionary lesson for our current preoccupation with genes. Eugenics was the effort to apply principles of genetics to improve the human race. Most people equate eugenics with the atrocities committed for the sake of racial purity in Nazi Germany. Most are unaware of the “positive” eugenics movement, exemplified in England, which advocated voluntary efforts by families to improve their own heredity. Fewer still realize that a coercive, “negative” eugenics movement flourished in the United States, that it involved numerous prominent scientists and civic lead- ers, and that it made its intellectual home at the forerunner of the now prestigious Cold Spring Harbor Laboratory. During the first decade of the 20th century, eugenics was organized as a scientific field by the con- fluence of Mendelian genetics and experimental breeding. This synthesis was embodied by Charles Benedict Davenport, who is considered the father of the American eugenics movement. When Charles Da v e n p o r t arrived at Cold Spring Harbor in 1898, he assumed the directorship of The Biological La b o r a t o r y, a prog r essive, if somewhat sleepy, “summer camp” for the study of evolution. -

The History of Neuroscience in Autobiography VOLUME 6 This Page Intentionally Left Blank the History of Neuroscience in Autobiography VOLUME 6
The History of Neuroscience in Autobiography VOLUME 6 This page intentionally left blank The History of Neuroscience in Autobiography VOLUME 6 Edited by Larry R. Squire 1 2009 1 Oxford University Press, Inc., publishes works that further Oxford University’s objective of excellence in research, scholarship, and education. Oxford New York Auckland Cape Town Dar es Salaam Hong Kong Karachi Kuala Lumpur Madrid Melbourne Mexico City Nairobi New Delhi Shanghai Taipei Toronto With offi ces in Argentina Austria Brazil Chile Czech Republic France Greece Guatemala Hungary Italy Japan Poland Portugal Singapore South Korea Switzerland Thailand Turkey Ukraine Vietnam Copyright © 2009 by Society for Neuroscience Published by Oxford University Press, Inc. 198 Madison Avenue, New York, New York 10016 www.oup.com Oxford is a registered trademark of Oxford University Press All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the prior permission of Oxford University Press. Library of Congress Control Number: 96070950 ISBN: 978–0–19–538010–1 1 3 5 7 9 8 6 4 2 Printed in the United States of America on acid-free paper Contents Previous Contributors vii Preface to Volume 1 ix Preface to Volume 6 xi Bernard W. Agranoff 1 Emilio Bizzi 32 Marian Cleeves Diamond 62 Charles G. Gross 96 Richard Held 158 Leslie L. Iversen 188 Masakazu Konishi 226 Lawrence Kruger 264 Susan E. Leeman 318 Vernon B. Mountcastle 342 Shigetada Nakanishi 382 Solomon H. Snyder 420 Nobuo Suga 480 Hans Thoenen 514 Index of Names This page intentionally left blank Previous Contributors Volume 1 Denise Albe-Fessard Herbert H.