Hydrostatic Skeletons in the Crustacea: Support During Molting in an Aquatic and a Terrestrial Crab
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chapter 14 1. Specialization of Regions of the Body for Specific
Chapter 14 1. Specialization of regions of the body for specific functions, as seen in arthropods, is called A) tagmatization. B) metamerism. C) truncation. D) differentiation. E) cephalization. 2. Members of class __________ are among the most numerous crustaceans, and are both marine and freshwater in distribution. A) Cirripedia B) Copepoda C) Branchiopoda D) Malacostraca E) Isopoda 3. Which type of crustacean do many zoologists believe to have the greatest number of individuals of any type of animal on the planet? A) isopods B) fairy shrimp C) brine shrimp D) copepods E) barnacles 4. Which of the following phyla of animals are not ecdysozoan? A) Arthropoda B) Nematoda C) Gastrotricha D) Nematomorpha E) Kinorhyncha 5. Which of the following is not a synapomorphy that unites the members of the ecdysozoan clade? A) the blastopore develops into the anus B) loss of epidermal cilia C) possession of a cuticle D) shedding of cuticle through ecdysis E) all of the above are synapomorphies shared by ecdysozoans Page 1 6. The __________ is the outer layer of the arthropod exoskeleton, and it is composed of a waterproofing waxy lipoprotein. A) lipocuticle B) mesocuticle C) epicuticle D) endocuticle E) sclerocuticle 7. The tough, leathery polysaccharide in the arthropod procuticle is A) lipoprotein. B) calcium carbonate. C) scleroprotein. D) chitin. E) glycogen. 8. The arthropod skeleton hardens by __________, which is a formation of chemical bonds between protein chains. A) carbonization B) tagmatization C) calcification D) chitinization E) sclerotization 9. Sensory receptors called __________ occur in the arthropod exoskeleton in the form of pegs, bristles, and lenses. -

Research Interests Related to the Cambridge-MIT Institute
Nanoscale Structural Design Principles of Biocomposite Exoskeletons As a Guide for New Energy-Absorbing Materials Technologies Team 1 : Energy Absorbing Materials : Multiscale Design and Evaluation of Nanostructured Materials for Ballistic and Blast Protection MIT Institute for Soldier Technologies (ISN) Christine Ortiz, Assistant Professor Massachusetts Institute of Technology, Department of Materials Science and Engineering WWW : http://web.mit.edu/cortiz/www/ August 2002 I. Background : Structural Design Principles and Energy Absorbing Mechanisms From millions of years of evolution, nature has ingeniously figured out innumerate structural design principles to produce multifunctional, and in many cases stimulus-responsive, materials with superior mechanical properties[1-6]. Examples of these include exoskeletons of many invertebrate animals such as mollusks, arthropods (e.g. crustaceans such as crabs, insects), cnidaria (e.g. corals), and structural components of mammals such as turtle shell, rhinocerous horn, bovine hoof horn, deer antlers, elephant tusks, and teeth. Most tough biological materials are complex, hierarchical, multilayered nanocomposites that undergo a wide variety of different energy-absorbing toughening mechanisms at many length scales. Some of these mechanisms in both biological and synthetic composite materials [7-10] are shown in Figure 1 and include; 1) rupture of "sacrificial" weaker bonds in the macromolecular component (e.g. Mollusk shell nacre), 2) extension, pull-out, and/or ligament formation of a macromolecular component bridging an interface (e.g. Mollusk shell nacre), 3) void formation (e.g. via cavitation of rubber particles in a thermoset composite or stress whitening in semicrystalline polymers) leading to bulk plastic deformation, crack blunting, pinning and branching, 4) localized plastic deformation ahead of a crack tip (e.g. -

A Review of Biological Fluid Power Systems and Their Potential Bionic Applications
The University of Manchester Research A review of biological fluid power systems and their potential bionic applications DOI: 10.1007/s42235-019-0031-6 Document Version Accepted author manuscript Link to publication record in Manchester Research Explorer Citation for published version (APA): Liu, C., Wang, Y., Ren, L., & Ren, L. (2019). A review of biological fluid power systems and their potential bionic applications. Journal of Bionic Engineering. https://doi.org/10.1007/s42235-019-0031-6 Published in: Journal of Bionic Engineering Citing this paper Please note that where the full-text provided on Manchester Research Explorer is the Author Accepted Manuscript or Proof version this may differ from the final Published version. If citing, it is advised that you check and use the publisher's definitive version. General rights Copyright and moral rights for the publications made accessible in the Research Explorer are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Takedown policy If you believe that this document breaches copyright please refer to the University of Manchester’s Takedown Procedures [http://man.ac.uk/04Y6Bo] or contact [email protected] providing relevant details, so we can investigate your claim. Download date:04. Oct. 2021 A review of biological fluid power systems and their potential bionic applications Chunbao Liu1,2, Yingjie Wang 1,2, Luquan Ren 2, Lei Ren2,3 1 School of Mechanical and Aerospace Engineering, Jilin University, Changchun 130022, China 2 Key Laboratory of Bionic Engineering, Ministry of Education, Jilin University, Changchun 130022, China 3 School of Mechanical, Aerospace and Civil Engineering, University of Manchester, Manchester M13 9PL, UK Abstract Nature has always inspired human achievements in industry, and biomimetics is increasingly being applied in fluid power technology. -

Chapter 1 and 2
CHAPTER 1 AND 2 Life is a unique, complex organization of molecules that expresses it self though chemical reactions which lead to growth, development, responsiveness, adaptation & reproduction. Unique features of living organesim: Growth- reproduction- metabolism- consciousness-life span. Living organisims are therefore, self- replicating, evolving & self-regulatory interactive systems capable of responding to external stimuli. Currently 1.7-1.8 billion living organisms known to science. Out of which 1.25 are animals and abut 0.5 millions are plants. Systematic is branch of biology that deals with cataloguing plants, animals and other organism into categories that can be named, compared & studied. Biology : father of biology- Aristotle,Biology (Bio-life form, logy-study) -Father of Zoology ( Aristotle ) Father of Botany ( Theophrastus ) Taxonomy: study of rules & procedure to classify organisms. Cell contains - Cytoplasm and Nucleoplasm Collectively called Protoplasm (Physical bas is of life ) given by purkinje Taxonomic categories (7 obligate ) - Kingdom (less similarities) - Phylum ( animal ) / Division (plant ) - Class - Order - Family - Genus - Species ( More similarity ) basic unit of classification Bionomical nomenclature given by C.linneaus Taxonomic AIDS: Herbarium : It is a place where dried and pressed specimens, mounted on sheets are kept systematically according bentham to bantams & hooker classification. It carries a label on right corner which provide. Information for future use. It provides a quick refer back system and is quite useful for people involved in taxonomic studies. All institutes leading with baotanical studies maintain their herbaria. HISTORY OF HERBARIA : Majority of the world’s famous herbaria originated from the botanical gardens. The first herbarium was set up at Pisa in Italy by a professor of botany Luca Ghini. -

P> TEST YOUR KNOWLEDGE Chapter 32: Introduction to Animal Evolution
TEST YOUR KNOWLEDGE Chapter 32: Introduction to Animal Evolution MULTIPLE CHOICE: Choose the one best answer. 1. Sponges differ from the rest of the animals be cause a. they are completely sessile. b. they have radial symmetry and are suspension- feeders. c -theirsimpIe-body structure rLas-notrue-nssues—-- and they have a unique embryology. d. they are not multicellular. e. they have no flagellated cells. 7. Which of the following is not descriptive of a 2.. An. insect larva pseudocoelomate? a. is a miniature version of the adult. a. a body cavity incompletely lined by mesoderm b. is transformed into an adult by molting. b. bilateral symmetry c. ensures more genetic variation in the insect life c. triploblastic cycle. d. true tissues d. is a sexually immature organism specialized e. schizocoelous formation of body cavity for eating and growth. e. is all of the above. 8. According to evidence from developmental genetics, the last common ancestor of the bilateral 3. Cephalization animals may have been a. is the development of bilateral symmetry. a. a parazoan that resembled a sponge. b. is the formation of a coelom by budding from b. a complex animal with a head, a longitudinal the archenteron. nerve cord, and a segmented body with legs. c. is a diagnostic characteristic of deuterostomes. c. a simple bilateral animal with no specializa d. is common in radially symmetrical animals. tion. e. is associated with motile animals that concen d. a colonial flagellated protist. trate sensory organs in a head region. e. a pseudocoelomate such as a roundworm. 4. A true coelom a. -

FISH310: Biology of Shellfishes
FISH310: Biology of Shellfishes Lecture Slides #3 Phylogeny and Taxonomy sorting organisms How do we classify animals? Taxonomy: naming Systematics: working out relationships among organisms Classification • All classification schemes are, in part, artificial to impose order (need to start some where using some information) – Cell number: • Acellular, One cell (_________), or More than one cell (metazoa) – Metazoa: multicellular, usu 2N, develop from blastula – Body Symmetry – Developmental Pattern (Embryology) – Evolutionary Relationship Animal Kingdom Eumetazoa: true animals Corals Anemones Parazoa: no tissues Body Symmetry • Radial symmetry • Phyla Cnidaria and Ctenophora • Known as Radiata • Any cut through center ! 2 ~ “mirror” pieces • Bilateral symmetry • Other phyla • Bilateria • Cut longitudinally to achieve mirror halves • Dorsal and ventral sides • Anterior and posterior ends • Cephalization and central nervous system • Left and right sides • Asymmetry uncommon (Porifera) Form and Life Style • The symmetry of an animal generally fits its lifestyle • Sessile or planktonic organisms often have radial symmetry • Highest survival when meet the environment equally well from all sides • Actively moving animals have bilateral symmetry • Head end is usually first to encounter food, danger, and other stimuli Developmental Pattern • Metazoa divided into two groups based on number of germ layers formed during embryogenesis – differs between radiata and bilateria • Diploblastic • Triploblastic Developmental Pattern.. • Radiata are diploblastic: two germ layers • Ectoderm, becomes the outer covering and, in some phyla, the central nervous system • Endoderm lines the developing digestive tube, or archenteron, becomes the lining of the digestive tract and organs derived from it, such as the liver and lungs of vertebrates Diploblastic http://faculty.mccfl.edu/rizkf/OCE1001/Images/cnidaria1.jpg Developmental Pattern…. -

CH33 Arthropods All.Pptx
10/15/14 Biosc 41 Announcements 10/15 Quick Platyhelminthes Review v What type of symmetry do animals in Phylum v Quiz today- Phyla Platyhelminthes and Mollusca Platyhelminthes have? v v Lecture- Phylum Arthropoda Digestive system: mouth only (protostomes) v How does gas exchange take place? v Lab- Phylum Arthropoda v What are protonephridia for? v What is the class of free-living flatworms? An v Mon’s lecture will finish any arthropod info we don’t get to example? today, have arthropod quiz, and do an exam review activity v What are the classes of parasitic flatworms? Example? v Mon’s lab will finish arthropod lab material (+ crayfish dissection) Quick Mollusca Review Quick Mollusca Review v Features and structures universal amongst molluscs: v Four major classes: § Mantle- secretes shell in shelled molluscs; sometimes § Class Polyplacophora- Chitons portions of mantle form siphon(s) § Class Gastropoda- snails, nudibranchs, slugs § Foot- contains sensory cells; adapted in various ways in § Life cycle includes trochophore & veliger larvae different classes § Veliger stage involves torsion § Visceral mass- internal organs consistent between classes, but arranged in different orientations § Class Bivalvia- clams, oysters, mussels § Life cycle includes trochophore & veliger larvae § Circulatory system includes heart and is open in all but cephalopods (which have a closed circulatory system) § Class Cephalopoda- squid, octopus, cuttlefish, nautilus § Nervous system includes nerve cords and centralized brain tissue § Mantle has been modified to include a siphon for locomotion § Digestive system includes radula, well-developed digestive gland, and “complete gut” § Foot has been modified into tentacles § Heart and kidneys function as excretory system § Closed circulation and complex brain Quick Mollusca Review Quiz 5 v What is a possible adverse effect of suspension feeding? 1. -
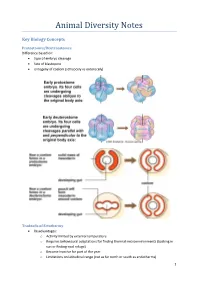
Animal Diversity Notes
Animal Diversity Notes Key Biology Concepts Protostomes/Deuterostomes Difference based on: type of embryo cleavage fate of blastopore ontogeny of coelom (schizocely vs enterocely) Tradeoffs of Ectothermy Disadvantages: o Activity limited by external temperature o Requires behavioural adaptations for finding thermal microenvironments (basking in sun or finding cool refuge) o Become inactive for part of the year o Limitations on latitudinal range (not as far north or south as endotherms) 1 Advantages: o High efficiency of converting ingested food to biomass o Can thrive in ecosystems of low productivity (deserts, hot and dry) o Can survive with low metabolic rate Purposes of Migration broadens the resource base, tracks available food high latitudes have longer days in summer, allowing extended foraging maintains relatively constant temperature by going tropics as winter sets in prevents permanent predation pressure in one location Advantages of the Coelomate Body Plan Cavity can function for circulation, waste disposal, and gamete storage and release Enables the development of a hydrostatic skeleton Provides cushioning for the digestive tract Muscular movements of the digestive tract isolated from the outer body wall and skeletal muscular movements Advantages of Metamerism Improved efficiency of motion using hydrostatic skeleton Independent nervous control and movement of segments Architectural redundancy allows specialisation of segments as well as survival/regeneration when segments are lost Modes of Development 2 3 Timeline of Evolution 4 Essential Diagrams Cladograms 5 6 7 8 9 10 Key Animal Diagrams 11 12 13 14 15 16 17 18 19 20 21 22 23 Animal Phyla Porifera Calcarea: calcareous sponges, mostly small Hexactinellida: possess six-rayed siliceous spicules, have continuous syncytial tissue called trabecular reticulum Demospongiae: Over 80% of sponge species, all are leuconoid, can have siliceous spicules or spongin fibres Cnidaria Hydrozoa: generally have asexual polyp and sexual medusa stages (e.g. -

The Crusty Fossils
Curriculum Units by Fellows of the Yale-New Haven Teachers Institute 1985 Volume VII: Skeletal Materials- Biomineralization The Crusty Fossils Curriculum Unit 85.07.03 by Carolyn C. Smith Stop! Look! Listen! These three basic commands can be associated with any adventure that a person wishes to undertake. As this unit is being presented those three little words will play a significant role. The changes which occur on this earth are so gradual that we don’t notice them immediately. However, if we were to ignore our surroundings for a few days or weeks, we would be able to recognize a difference of some kind almost instantly. When we finally notice the changes we are quick to say, “It happened overnight!” Scientists are constantly recording the changes which are occurring on this earth. For centuries man has theorized why, when, where, who, what, and how something is done. Ever so often new evidence is presented and he is forced to change his explanations when results contradict his previous understanding. This unit deals with the results of a chain of events which is believed to have started 4-1/2 billion years ago. We are going to take a look at a species of animals which is found in the Arthropoda Kingdom—“The Crab”. You will find that these little creatures are quite fascinating in their developmental and behavior patterns. After studying this unit the students will be able to: 1. identify the Phylum Arthropoda. 2. discuss the Class Crustacea. 3. discuss the evolution of the crabs. 4. discuss the stages of development of crabs. -

A Closer Look at Arthropods 701 DO NOT EDIT--Changes Must Be Made Through “File Info” Correctionkey=A
DO NOT EDIT--Changes must be made through “File info” CorrectionKey=A A Closer Look CHAPTER 24 at Arthropods BIg IdEa Arthropods are invertebrates and are the most abundant and diverse of all animals. 24.1 Arthropod diversity 7A, 7E, 8C 24.2 Crustaceans 7A, 7E, 8C 24.3 Arachnids 8C data analysis Constructing SCatterplots 2F, 2G 24.4 Insect adaptations 24.5 Arthropods and Humans Online BiOlOgy HMDScience.com ONLINE Labs ■■ Virtual Lab Insects and Crime Scene Analysis ■■ QuickLab Comparing Arthropods ■■ Video Lab Butterfly Metamorphosis ■■ Hatching Brine Shrimp ■■ S.T.E.M. Lab Exoskeleton Strength ■■ Daphnia and Heart Rate ■■ Inside a Crayfish ■■ Identifying Arthropods in a Decomposer System ■■ Determining Time of Death Using Entomology (t) Tenaglia-missouriplants.com ©Dan 700 Unit 8: Animals DO NOT EDIT--Changes must be made through “File info” CorrectionKey=A Q What is the relationship between these two insects? Arthropod predators such as this digger wasp help to keep an important balance among Earth’s invertebrates. This digger wasp has captured a meal, not for itself but for its young. The wasp will deposit the live, but paralyzed, grasshopper into a burrow she has constructed. She will then lay a single egg next to the grasshopper so when the egg hatches the larva will have a fresh meal. r E a d IN g T o o lb o x This reading tool can help you learn the material in the following pages. uSINg LaNGUAGE YOur TurN Classification Categories are groups of things that Read the following sentences. Identify the category and have certain characteristics in common. -

Ultrastructure, Histochemistry, and Mineralization Patterns in the Ecdysial Suture of the Blue Crab, Callinectes Sapidus
ULTRASTRUCTURE, HISTOCHEMISTRY, AND MINERALIZATION PATTERNS IN THE ECDYSIAL SUTURE OF THE BLUE CRAB, CALLINECTES SAPIDUS Carolina Priester A Thesis Submitted to the University of North Carolina at Wilmington in Partial Fulfillment of the Requirements for the Degree of Master of Science Department of Biological Sciences University of North Carolina at Wilmington 2003 Approved by Advisory Committee Neil Hadley Robert Roer Dr. Neil Hadley Dr. Robert Roer Thomas H. Shafer Richard M. Dillaman Dr. Thomas Shafer Dr. Richard Dillaman, Chair Accepted by Robert Roer Dean, Graduate School This thesis has been prepared in the style and format consistent with the journal Journal of Morphology ii TABLE OF CONTENTS ABSTRACT....................................................................................................................... iv ACKNOWLEDGEMENTS............................................................................................... vi DEDICATION.................................................................................................................. vii LIST OF TABLES........................................................................................................... viii LIST OF FIGURES ........................................................................................................... ix INTRODUCTION ...............................................................................................................1 MATERIALS AND METHODS.........................................................................................5 -

Glossary.Pdf
Glossary Pronunciation Key accessory fruit A fruit, or assemblage of fruits, adaptation Inherited characteristic of an organ- Pronounce in which the fleshy parts are derived largely or ism that enhances its survival and reproduc- a- as in ace entirely from tissues other than the ovary. tion in a specific environment. – Glossary Ј Ј a/ah ash acclimatization (uh-klı¯ -muh-tı¯-za -shun) adaptive immunity A vertebrate-specific Physiological adjustment to a change in an defense that is mediated by B lymphocytes ch chose environmental factor. (B cells) and T lymphocytes (T cells). It e¯ meet acetyl CoA Acetyl coenzyme A; the entry com- exhibits specificity, memory, and self-nonself e/eh bet pound for the citric acid cycle in cellular respi- recognition. Also called acquired immunity. g game ration, formed from a fragment of pyruvate adaptive radiation Period of evolutionary change ı¯ ice attached to a coenzyme. in which groups of organisms form many new i hit acetylcholine (asЈ-uh-til-ko–Ј-le¯n) One of the species whose adaptations allow them to fill dif- ks box most common neurotransmitters; functions by ferent ecological roles in their communities. kw quick binding to receptors and altering the perme- addition rule A rule of probability stating that ng song ability of the postsynaptic membrane to specific the probability of any one of two or more mu- o- robe ions, either depolarizing or hyperpolarizing the tually exclusive events occurring can be deter- membrane. mined by adding their individual probabilities. o ox acid A substance that increases the hydrogen ion adenosine triphosphate See ATP (adenosine oy boy concentration of a solution.