The Nature of Ordovician Limestone-Marl Alternations in the Oslo-Asker District
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Permo-Carboniferous Oslo Rift Through Six Stages and 65 Million Years
52 by Bjørn T. Larsen1, Snorre Olaussen2, Bjørn Sundvoll3, and Michel Heeremans4 The Permo-Carboniferous Oslo Rift through six stages and 65 million years 1 Det Norske Oljeselskp ASA, Norway. E-mail: [email protected] 2 Eni Norge AS. E-mail: [email protected] 3 NHM, UiO. E-mail: [email protected] 4 Inst. for Geofag, UiO. E-mail: [email protected] The Oslo Rift is the northernmost part of the Rotliegen- des basin system in Europe. The rift was formed by lithospheric stretching north of the Tornquist fault sys- tem and is related tectonically and in time to the last phase of the Variscan orogeny. The main graben form- ing period in the Oslo Region began in Late Carbonif- erous, culminating some 20–30 Ma later with extensive volcanism and rifting, and later with uplift and emplacement of major batholiths. It ended with a final termination of intrusions in the Early Triassic, some 65 Ma after the tectonic and magmatic onset. We divide the geological development of the rift into six stages. Sediments, even with marine incursions occur exclusively during the forerunner to rifting. The mag- matic products in the Oslo Rift vary in composition and are unevenly distributed through the six stages along the length of the structure. Introduction The Oslo Palaeorift (Figure 1) contributed to the onset of a pro- longed period of extensional faulting and volcanism in NW Europe, which lasted throughout the Late Palaeozoic and the Mesozoic eras. Widespread rifting and magmatism developed north of the foreland of the Variscan Orogen during the latest Carboniferous and contin- ued in some of the areas, like the Oslo Rift, all through the Permian period. -

Early Silurian Oceanic Episodes and Events
Journal of the Geological Society, London, Vol. 150, 1993, pp. 501-513, 3 figs. Printed in Northern Ireland Early Silurian oceanic episodes and events R. J. ALDRIDGE l, L. JEPPSSON 2 & K. J. DORNING 3 1Department of Geology, The University, Leicester LE1 7RH, UK 2Department of Historical Geology and Palaeontology, SiSlvegatan 13, S-223 62 Lund, Sweden 3pallab Research, 58 Robertson Road, Sheffield $6 5DX, UK Abstract: Biotic cycles in the early Silurian correlate broadly with postulated sea-level changes, but are better explained by a model that involves episodic changes in oceanic state. Primo episodes were characterized by cool high-latitude climates, cold oceanic bottom waters, and high nutrient supply which supported abundant and diverse planktonic communities. Secundo episodes were characterized by warmer high-latitude climates, salinity-dense oceanic bottom waters, low diversity planktonic communities, and carbonate formation in shallow waters. Extinction events occurred between primo and secundo episodes, with stepwise extinctions of taxa reflecting fluctuating conditions during the transition period. The pattern of turnover shown by conodont faunas, together with sedimentological information and data from other fossil groups, permit the identification of two cycles in the Llandovery to earliest Weniock interval. The episodes and events within these cycles are named: the Spirodden Secundo episode, the Jong Primo episode, the Sandvika event, the Malm#ykalven Secundo episode, the Snipklint Primo episode, and the lreviken event. Oceanic and climatic cyclicity is being increasingly semblages (Johnson et al. 1991b, p. 145). Using this recognized in the geological record, and linked to major and approach, they were able to detect four cycles within the minor sedimentological and biotic fluctuations. -

Agenda 2030 in Asker
Agenda 2030 in Asker Voluntary local review 2021 Content Opening Statement by mayor Lene Conradi ....................................4 Highlights........................................................................................5 Introduction ....................................................................................6 Methodology and process for implementing the SDGs ...................8 Incorporation of the Sustainable Development Goals in local and regional frameworks ........................................................8 Institutional mechanisms for sustainable governance ....................... 11 Practical examples ........................................................................20 Sustainability pilots .........................................................................20 FutureBuilt, a collaboration for sustainable buildings and arenas .......20 Model projects in Asker ...................................................................20 Citizenship – evolving as a co-creation municipality ..........................24 Democratic innovation.....................................................................24 Arenas for co-creation and community work ....................................24 Policy and enabling environment ..................................................26 Engagement with the national government on SDG implementation ...26 Cooperation across municipalities and regions ................................26 Creating ownership of the Sustainable Development Goals and the VLR .......................................................................... -
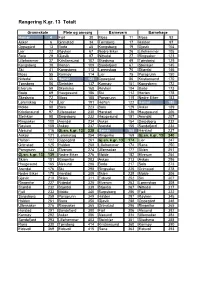
Rangering K.Gr. 13 Totalt
Rangering K.gr. 13 Totalt Grunnskole Pleie og omsorg Barnevern Barnehage Hamar 4 Fjell 30 Moss 11 Moss 92 Asker 6 Grimstad 34 Tønsberg 17 Halden 97 Oppegård 13 Bodø 45 Kongsberg 19 Gjøvik 104 Lier 22 Røyken 67 Nedre Eiker 26 Lillehammer 105 Sola 29 Gjøvik 97 Nittedal 27 Ringsaker 123 Lillehammer 37 Kristiansund 107 Skedsmo 49 Tønsberg 129 Kongsberg 38 Horten 109 Sandefjord 67 Steinkjer 145 Ski 41 Kongsberg 113 Lørenskog 70 Stjørdal 146 Moss 55 Karmøy 114 Lier 75 Porsgrunn 150 Nittedal 55 Hamar 123 Oppegård 86 Kristiansund 170 Tønsberg 56 Steinkjer 137 Karmøy 101 Kongsberg 172 Elverum 59 Skedsmo 168 Røyken 104 Bodø 173 Bodø 69 Haugesund 186 Ski 112 Horten 178 Skedsmo 72 Moss 188 Porsgrunn 115 Nedre Eiker 183 Lørenskog 74 Lier 191 Horten 122 Hamar 185 Molde 88 Sola 223 Sola 129 Asker 189 Kristiansund 97 Ullensaker 230 Harstad 136 Haugesund 206 Steinkjer 98 Sarpsborg 232 Haugesund 151 Arendal 207 Ringsaker 100 Arendal 234 Asker 154 Sarpsborg 232 Røyken 108 Askøy 237 Arendal 155 Sandefjord 234 Ålesund 116 Gj.sn. k.gr. 13 238 Hamar 168 Harstad 237 Askøy 121 Lørenskog 254 Ringerike 169 Gj.sn. k.gr. 13 240 Horten 122 Oppegård 261 Gj.sn. k.gr. 13 174 Lier 247 Grimstad 125 Halden 268 Lillehammer 174 Rana 250 Porsgrunn 133 Elverum 274 Ullensaker 177 Skien 251 Gj.sn. k.gr. 13 139 Nedre Eiker 276 Molde 182 Elverum 254 Skien 151 Ringerike 283 Askøy 213 Askøy 256 Haugesund 165 Ålesund 288 Bodø 217 Sola 273 Arendal 176 Ski 298 Ringsaker 225 Grimstad 278 Nedre Eiker 179 Harstad 309 Skien 239 Molde 306 Gjøvik 210 Skien 311 Eidsvoll 252 Ski 307 Ringerike -

Camilla Brautaset 26 Petra Hyncicova 27 Ane Johnsen 28 Jesse Knori 29-30 Anne Siri Lervik 31 Lucy Newman 32-33 Christina Rolandsen 34
Women’s Nordic Camilla Brautaset 26 Petra Hyncicova 27 Ane Johnsen 28 Jesse Knori 29-30 Anne Siri Lervik 31 Lucy Newman 32-33 Christina Rolandsen 34 25 2017 colorado buffaloes Camilla Brautaset A A A 5-5 Senior Women’s Nordic Oslo, Norway (Oslo Handelsgym/Heming Ski Club) 3 Letters; 2014 as a freshman, 2015 as a sophomore, 2016 as a junior Career at Colorado— 2016 (Junior)— An experienced veteran, she enters her senior campaign with 30 races under her belt - the most by any Buffalo on the women’s Nordic team entering the 2017 season. SEASON BY SEASON RESULTS She finished all 10 races through the RMISA Championships and had seven top 20 finishes. 2014 CL FS She placed ninth in the classic race at the RMISA Championships for her top finish of the season in her very last meet. Her top freestyle result was 12th place in the freestyle at New 2M0e1x5ic o(.S Sohpeh womaso ar me)e— mber of the National Collegiate All-Academic Ski Team for maintaining above a 3.5 grade point average and participating at the RMISA Championships/NCAA West Regional. Utah Invitational ̶̶ Montana State Invitational 8 11 She competed and finished 10 events, with four top 20 and two top Colorado Invitational 23 11 2150 1ap4p (eFarreasnhcmesa. Ant) —the New Mexico Invitational, she placed tenth in the 10k classic. She placed eleventh in the 10k classic at the Utah Invitational. She was a member of CU’s 4.0 Club and is N20ew15 Mexico Invitational C45L FS a part of the National All Academic Ski Team. -

Supplementary File for the Paper COVID-19 Among Bartenders And
Supplementary file for the paper COVID-19 among bartenders and waiters before and after pub lockdown By Methi et al., 2021 Supplementary Table A: Overview of local restrictions p. 2-3 Supplementary Figure A: Estimated rates of confirmed COVID-19 for bartenders p. 4 Supplementary Figure B: Estimated rates of confirmed COVID-19 for waiters p. 4 1 Supplementary Table A: Overview of local restrictions by municipality, type of restriction (1 = no local restrictions; 2 = partial ban; 3 = full ban) and week of implementation. Municipalities with no ban (1) was randomly assigned a hypothetical week of implementation (in parentheses) to allow us to use them as a comparison group. Municipality Restriction type Week Aremark 1 (46) Asker 3 46 Aurskog-Høland 2 46 Bergen 2 45 Bærum 3 46 Drammen 3 46 Eidsvoll 1 (46) Enebakk 3 46 Flesberg 1 (46) Flå 1 (49) Fredrikstad 2 49 Frogn 2 46 Gjerdrum 1 (46) Gol 1 (46) Halden 1 (46) Hemsedal 1 (52) Hol 2 52 Hole 1 (46) Hurdal 1 (46) Hvaler 2 49 Indre Østfold 1 (46) Jevnaker1 2 46 Kongsberg 3 52 Kristiansand 1 (46) Krødsherad 1 (46) Lier 2 46 Lillestrøm 3 46 Lunner 2 46 Lørenskog 3 46 Marker 1 (45) Modum 2 46 Moss 3 49 Nannestad 1 (49) Nes 1 (46) Nesbyen 1 (49) Nesodden 1 (52) Nittedal 2 46 Nordre Follo2 3 46 Nore og Uvdal 1 (49) 2 Oslo 3 46 Rakkestad 1 (46) Ringerike 3 52 Rollag 1 (52) Rælingen 3 46 Råde 1 (46) Sarpsborg 2 49 Sigdal3 2 46 Skiptvet 1 (51) Stavanger 1 (46) Trondheim 2 52 Ullensaker 1 (52) Vestby 1 (46) Våler 1 (46) Øvre Eiker 2 51 Ål 1 (46) Ås 2 46 Note: The random assignment was conducted so that the share of municipalities with ban ( 2 and 3) within each implementation weeks was similar to the share of municipalities without ban (1) within the same (actual) implementation weeks. -

Upcoming Projects Infrastructure Construction Division About Bane NOR Bane NOR Is a State-Owned Company Respon- Sible for the National Railway Infrastructure
1 Upcoming projects Infrastructure Construction Division About Bane NOR Bane NOR is a state-owned company respon- sible for the national railway infrastructure. Our mission is to ensure accessible railway infra- structure and efficient and user-friendly ser- vices, including the development of hubs and goods terminals. The company’s main responsible are: • Planning, development, administration, operation and maintenance of the national railway network • Traffic management • Administration and development of railway property Bane NOR has approximately 4,500 employees and the head office is based in Oslo, Norway. All plans and figures in this folder are preliminary and may be subject for change. 3 Never has more money been invested in Norwegian railway infrastructure. The InterCity rollout as described in this folder consists of several projects. These investments create great value for all travelers. In the coming years, departures will be more frequent, with reduced travel time within the InterCity operating area. We are living in an exciting and changing infrastructure environment, with a high activity level. Over the next three years Bane NOR plans to introduce contracts relating to a large number of mega projects to the market. Investment will continue until the InterCity rollout is completed as planned in 2034. Additionally, Bane NOR plans together with The Norwegian Public Roads Administration, to build a safer and faster rail and road system between Arna and Stanghelle on the Bergen Line (western part of Norway). We rely on close -

(Mecoptera) in Norway
© Norwegian Journal of Entomology. 21 June 2011 Distribution of Boreus westwoodi Hagen, 1866 and Boreus hyemalis (L., 1767) (Mecoptera) in Norway SIGMUND HÅGVAR & EIVIND ØSTBYE Hågvar, S. & Østbye, E. 2011. Distribution of Boreus westwoodi Hagen, 1866 and Boreus hyemalis (L., 1767) (Mecoptera) in Norway. Norwegian Journal of Entomology 58, 73–80. An extensive material collected during nearly fifty years adds new detailed information on the distribution of the winter active insects Boreus westwoodi Hagen, 1866 and B. hyemalis (L., 1767) in Norway. Since females are difficult to identify, the new data rely on males. Based on the revised Strand-system, the following geographical regions are new to B. westwoodi: Ø, BØ, VAY, ON, TEI, TEY, MRI, MRY, and TRY. For B. hyemalis, AK, BØ, TEI, RY, SFI, and NTI are new regions. While B. westwoodi is widespread in Norway, including the three northernmost counties, B. hyemalis seems to be restricted to the south, with the northernmost record in NTI. In Sweden, the situation is similar: B. westwoodi is widespread, while B. hyemalis has been recorded as far north as Västerbotten, at a latitude corresponding to the northernmost record in Norway. The known distribution of both species in Norway is presented on EIS-grid map. Key words: Boreus hyemalis, Boreus westwoodi, Mecoptera, distribution, Norway. Sigmund Hågvar, Department of Ecology and Natural Resource Management, P.O. Box 5003, Norwegian University of Life Sciences, NO-1432 Ås, Norway. E-mail: [email protected] Eivind Østbye, Ringeriksveien 580, NO-3410 Sylling, Norway. E-mail: [email protected] Introduction county was described by Greve (1966). -

LISTE OVER TROSSAMFUNN I BUSKERUD Pr
LISTE OVER TROSSAMFUNN I BUSKERUD pr. 01.01.2017 Adresse Besøksadr. Forstander Vigsels- Org. Nr.: myndighet Navn Islam: Afghaneres kulturelle og Øvre Eikervei 75, 3048 Safi Mashukulla 990889880 Islamiske forening i Drammen Buskerud Anjuman-E-Islahul Tordenskioldsgt. 86, Anjem, Abdul Rehman 974 256 258 Muslimeen of Drammen 3044 Drammen Norway Anjumane-Islah-Ul Lier c/o Asif Rana, Asif Rana 889 585 692 Svenskerud 81, 3408 Tranby Buskerud og Vestfold Postboks 2011, 3003 Tollbugata 12, Ismail Yusuf Mohammed- 985 663 882 muslimsk trossamfunnet Drammen Drammen Adur Den Allevitiske Konnerudgata 31, 3045 Ali Ihsan Pervane 885 307 612 Trossamfunn i Norge Drammen Den Islamske Kurdiske c/o Abdul Rahman Shaw Ibrahim Salih 989631896 Forening i Drammen Hussein, Lierstranda 89, 3400 Lier Det afghanske kultur og c/o Suhailla Issa Boks Suhailla Issa 991231099 trossamfunn i Norge 9202, 3028 Drammen Det albansk kultur og Engene 70, Abedin Osmani 987436441 trossamfunn i Norge 3015 Drammen Asselam Center (Det c/o Hussam Algazban, Hussam Algazban 992195401 irakiske kultur og Åslyveien 27, trossamfunn) 3023 Drammen Det Islamske Kultur Senter i Postboks 2435, Colletsgt. 10, Ali Ekiz V 971 307 323 Drammen 3003 Drammen Drammen Det Islamske Kultursenter i Gamle Riksvei 242 Ilyas Tuzkaya 980 764 249 Nedre Eiker 3055 Krokstadelva Det Islamske forbundet i Nordahl Brunsgate 1, Nasseraldeen Saleh 994 989 197 Buskerud 3018 Drammen Det Tyrkiske Trossamfunn i Postboks 9705 Rømersvei 4, Orhan Al V 987 751 142 Drammen og Omegn 3010 Drammen Drammen Drammen Tyrkiske Tollbugt.39, Mehmet Beles 993 813 303 Islamske Menighet 3044 Drammen Hamwatan Islamsk og c/o Mirpadesha Steinbergvn 2 Mirpadesha Kohdamani, 998870593 Kulturell forening Kohdamani, boks 600 3050 Mjøndalen Coop Mega, Berja, 3605 Kongsberg Hallingdal Islamsk Senter Sentrumvegen 67, 3550 Abdifatah Isak Hassan 998 659 485 Gol Hønefoss islamsk senter Blomsgt. -

16Th General Report on the CPT's Activities Covering the Period 1 August 2005 to 31 July 2006
CPT/Inf (2006) 35 European Committee for the Prevention of Torture and Inhuman or Degrading Treatment or Punishment (CPT) 16th General Report on the CPT's activities covering the period 1 August 2005 to 31 July 2006 Strasbourg, 16 October 2006 The CPT is required to draw up every year a general report on its activities, which is published. This 16th General Report, as well as previous general reports and other information about the work of the CPT, may be obtained from the Committee's Secretariat or from its website: Secretariat of the CPT Human Rights Building Council of Europe F-67075 Strasbourg Cedex, France Tel: +33 (0)3 88 41 39 39 Fax: +33 (0)3 88 41 27 72 E-mail: [email protected] Web: http://www.cpt.coe.int Database: http://hudoc.cpt.coe.int CPT: 16TH GENERAL REPORT3 TABLE OF CONTENTS Page PREFACE................................................................................................................................................................5 ACTIVITIES DURING THE PERIOD 1 AUGUST 2005 TO 31 JULY 2006................................................7 Visits.........................................................................................................................................................7 Meetings and working methods..............................................................................................................10 Publications ............................................................................................................................................11 ORGANISATIONAL MATTERS......................................................................................................................12 -

Uttalelse Til Forslag Til Detaljregulering for Lloyds Marked I Hønefoss
Vår dato: Vår ref: 30.08.2021 2021/15632 Deres dato: Deres ref: 04.06.2021 20/10633 Ringerike kommune Saksbehandler, innvalgstelefon Postboks 123 Sentrum Brede Kihle, 32266865 3502 HØNEFOSS Ringerike kommune - Uttalelse til forslag til detaljregulering for Lloyds marked i Hønefoss Vi viser til brev av 4. juni 2021 med forslag til detaljregulering for Lloyds marked. Bakgrunn Det fremgår av oversendelsen at hensikten med planarbeidet er å legge til rette for ny bebyggelse i form av forretninger, tjenesteyting, kontor og hotell. Området ligger sentrumsnært ved Hønefossen i et attraktivt område av byen. Det tidligere industriområdet vil bli transformert til et mere moderne næringsområde. Ny bebyggelse skal utformes i kontrast til det gamle, samtidig som det skal sikres et helhetlig kulturmiljø med god balanse mellom gammelt og nytt. Bygninger med høy symbol- eller identitetsverdi skal bevares. Forslaget er utformet i tråd med en mulighetsstudie for området utført av arkitektfirmaet Snøhetta i 2019. Videre bygger detaljreguleringen på områdereguleringen for Hønefoss som ble vedtatt i 5. september 2019. Sentralt i området er det blant annet foreslått et bygg på 14 etasjer omtalt som Tårnet. Arealet her er foreslått regulert til hotell og kontor. Det er ikke foreslått boliger innenfor området. Vi ga innspill til planarbeidet i vårt brev av 28. juli 2016. Statsforvalterens rolle Vi skal bidra til at planer ivaretar nasjonale og vesentlige regionale interesser innen landbruk, klima og miljøvern, folkehelse, barn og unges interesser, samfunnssikkerhet og gravplasser. Statsforvalteren skal arbeide for at Stortingets og regjeringens vedtak, mål og retningslinjer innen våre ansvarsområder blir fulgt opp i kommunale planer. Kommunen er planmyndighet og har ansvaret for at plan- og bygningslovens formelle krav til innhold og planprosess oppfylles i planarbeidet. -

Lions Clubs International Club Membership Register Summary the Clubs and Membership Figures Reflect Changes As of September 2004
LIONS CLUBS INTERNATIONAL CLUB MEMBERSHIP REGISTER SUMMARY THE CLUBS AND MEMBERSHIP FIGURES REFLECT CHANGES AS OF SEPTEMBER 2004 CLUB CLUB LAST MMR FCL YR MEMBERSHI P CHANGES TOTAL DIST IDENT NBR CLUB NAME STATUS RPT DATE OB NEW RENST TRANS DROPS NETCG MEMBERS 3929 019603 AL 104 G 4 09-2004 32 0 0 0 0 0 32 3929 019607 DRAMMEN 104 G 4 08-2004 17 0 0 0 0 0 17 3929 019608 DRAMMEN VEST 104 G 4 07-2004 29 0 0 0 -1 -1 28 3929 019611 FLA 104 G 4 16 0 0 0 0 0 16 3929 019613 KRODSHERAD 104 G 4 09-2004 20 0 0 0 -1 -1 19 3929 019614 GOL 104 G 4 09-2004 23 1 0 0 0 1 24 3929 019616 HONEFOSS 104 G 4 27 0 0 0 0 0 27 3929 019617 HOL 104 G 4 09-2004 30 0 0 0 -1 -1 29 3929 019618 HURUM 104 G 4 08-2004 18 0 0 0 0 0 18 3929 019619 KONGSBERG 104 G 4 09-2004 36 1 0 0 -1 0 36 3929 019624 LIER 104 G 4 08-2004 27 0 0 0 -1 -1 26 3929 019625 LIER OST 104 G 4 24 0 0 0 0 0 24 3929 019629 MODUM 104 G 4 07-2004 28 0 0 0 -1 -1 27 3929 019631 NEDRE EIKER L C 104 G 4 04-2004 34 0 0 0 0 0 34 3929 019634 OVRE EIKER L C 104 G 4 09-2004 37 3 0 0 0 3 40 3929 019639 ROYKEN 104 G 4 09-2004 21 1 0 0 0 1 22 3929 019640 SIGDAL 104 G 4 09-2004 37 0 0 0 0 0 37 3929 019646 TYRISTRAND 104 G 4 09-2004 28 0 0 0 0 0 28 3929 019649 ASKER 104 G 4 09-2004 31 0 0 0 -1 -1 30 3929 019650 BORGEN 104 G 4 09-2004 11 0 0 0 0 0 11 3929 019652 BLAKSTAD 104 G 4 09-2004 22 0 0 0 -2 -2 20 3929 019663 NESBRU 104 G 4 08-2004 26 0 0 0 0 0 26 3929 019664 NESOYA 104 G 4 17 0 0 0 0 0 17 3929 019707 HOLMESTRAND 104 G 4 09-2004 30 0 0 0 -1 -1 29 3929 019709 HORTEN 104 G 4 09-2004 26 0 0 0 -2 -2 24 3929 029133