Early Silurian Oceanic Episodes and Events
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CONODONT BIOSTRATIGRAPHY and ... -.: Palaeontologia Polonica
CONODONT BIOSTRATIGRAPHY AND PALEOECOLOGY OF THE PERTH LIMESTONE MEMBER, STAUNTON FORMATION (PENNSYLVANIAN) OF THE ILLINOIS BASIN, U.S.A. CARl B. REXROAD. lEWIS M. BROWN. JOE DEVERA. and REBECCA J. SUMAN Rexroad , c.. Brown . L.. Devera, 1.. and Suman, R. 1998. Conodont biostrati graph y and paleoec ology of the Perth Limestone Member. Staunt on Form ation (Pennsy lvanian) of the Illinois Basin. U.S.A. Ill: H. Szaniawski (ed .), Proceedings of the Sixth European Conodont Symposium (ECOS VI). - Palaeont ologia Polonica, 58 . 247-259. Th e Perth Limestone Member of the Staunton Formation in the southeastern part of the Illinois Basin co nsists ofargill aceous limestone s that are in a facies relati on ship with shales and sandstones that commonly are ca lcareous and fossiliferous. Th e Perth conodo nts are do minated by Idiognathodus incurvus. Hindeodus minutus and Neognathodu s bothrops eac h comprises slightly less than 10% of the fauna. Th e other spec ies are minor consti tuents. The Perth is ass igned to the Neog nathodus bothrops- N. bassleri Sub zon e of the N. bothrops Zo ne. but we were unable to co nfirm its assignment to earliest Desmoin esian as oppose d to latest Atokan. Co nodo nt biofacies associations of the Perth refle ct a shallow near- shore marine environment of generally low to moderate energy. but locali zed areas are more variable. particul ar ly in regard to salinity. K e y w o r d s : Co nodo nta. biozonation. paleoecology. Desmoinesian , Penn sylvanian. Illinois Basin. U.S.A. -

Conodonts in Ordovician Biostratigraphy
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Archivio istituzionale della ricerca - Università di Modena e Reggio Emilia 1 Conodonts in Ordovician biostratigraphy STIG M. BERGSTRÖM AND ANNALISA FERRETTI Conodonts in Ordovician biostratigraphy The long time interval after Pander’s (1856) original conodont study can in terms of Ordovician conodont biostratigraphic research be subdivided into three periods, namely the Pioneer Period (1856-1955), the Transition Period (1955-1971), and the Modern Period (1971-Recent). During the pre-1920s, the few published conodont investigations were restricted to Europe and North America and were not concerned about the potential use of conodonts as guide fossils. Although primarily of taxonomic nature, the pioneer studies by Branson & Mehl, Stauffer, and Furnish during the 1930s represent the beginning of the use of conodonts in Ordovician biostratigraphy. However, no formal zones were introduced until Lindström (1955) proposed four conodont zones in the Lower Ordovician of Sweden, which marks the end of the Pioneer Period. Because Lindström’s zone classification was not followed by similar work outside Baltoscandia, the time interval up to the late 1960s can be regarded as a Transition Period. A milestone symposium volume, entitled ‘Conodont Biostratigraphy’ and published in 1971, 2 summarized much new information on Ordovician conodont biostratigraphy and is taken as the beginning of the Modern Period of Ordovician conodont biostratigraphy. In this volume, the Baltoscandic Ordovician was subdivided into named conodont zones whereas the North American Ordovician succession was classified into a series of lettered or numbered Faunas. Although most of the latter did not receive zone names until 1984, this classification has been used widely in North America. -

Revised Correlation of Silurian Provincial Series of North America with Global and Regional Chronostratigraphic Units 13 and D Ccarb Chemostratigraphy
Revised correlation of Silurian Provincial Series of North America with global and regional chronostratigraphic units 13 and d Ccarb chemostratigraphy BRADLEY D. CRAMER, CARLTON E. BRETT, MICHAEL J. MELCHIN, PEEP MA¨ NNIK, MARK A. KLEFF- NER, PATRICK I. MCLAUGHLIN, DAVID K. LOYDELL, AXEL MUNNECKE, LENNART JEPPSSON, CARLO CORRADINI, FRANK R. BRUNTON AND MATTHEW R. SALTZMAN Cramer, B.D., Brett, C.E., Melchin, M.J., Ma¨nnik, P., Kleffner, M.A., McLaughlin, P.I., Loydell, D.K., Munnecke, A., Jeppsson, L., Corradini, C., Brunton, F.R. & Saltzman, M.R. 2011: Revised correlation of Silurian Provincial Series of North America with global 13 and regional chronostratigraphic units and d Ccarb chemostratigraphy. Lethaia,Vol.44, pp. 185–202. Recent revisions to the biostratigraphic and chronostratigraphic assignment of strata from the type area of the Niagaran Provincial Series (a regional chronostratigraphic unit) have demonstrated the need to revise the chronostratigraphic correlation of the Silurian System of North America. Recently, the working group to restudy the base of the Wen- lock Series has developed an extremely high-resolution global chronostratigraphy for the Telychian and Sheinwoodian stages by integrating graptolite and conodont biostratigra- 13 phy with carbonate carbon isotope (d Ccarb) chemostratigraphy. This improved global chronostratigraphy has required such significant chronostratigraphic revisions to the North American succession that much of the Silurian System in North America is cur- rently in a state of flux and needs further refinement. This report serves as an update of the progress on recalibrating the global chronostratigraphic correlation of North Ameri- can Provincial Series and Stage boundaries in their type area. -

A New Species of the Conodont Genus Siphonodella Branson & Mehl
Estonian Journal of Earth Sciences, 2017, 66, 4, 188–192 https://doi.org/10.3176/earth.2017.15 A new species of the conodont genus Siphonodella Branson & Mehl (late Tournaisian) Andrey V. Zhuravlev Institute of Geology Komi SC, UrB RAS, Pervomayskaya 54, 167000 Syktyvkar, Russia; [email protected] Received 3 April 2017, accepted 6 June 2017, available online 16 October 2017 Abstract. A new upper Tournaisian (Lower Carboniferous) siphonodellid conodont species Siphonodella carinata n. sp. is described. The material comes from the shallow-water carbonate sediments of the Pechora Swell (Timan-Pechora region or NE of European Russia). The co-occurrence of conodonts Hindeodus cristulus (Youngquist & Miller), Bispathodus stabilis (Branson & Mehl) Morphotype 1, Polygnathus longiposticus Branson & Mehl and Pseudopolygnathus nodomarginatus (Branson) suggests the late Tournaisian (Lower Siphonodella crenulata Zone) age of the deposits. Morphologically the new species is similar to Siphonodella semichatovae Kononova & Lipnjagov and S. ludmilae Zhuravlev & Plotitsyn, but differs in possessing three rostral ridges at the late stages of ontogeny and Class III symmetry. The presence of the shallow-water siphonodellids Siphonodella bella Kononova & Migdisova and S. quasinuda Gagiev, Kononova & Pazuhin in the upper part of the Tournaisian is detected for the first time. Key words: Conodonta, new species, Siphonodella carinata n. sp., Lower Carboniferous, Tournaisian. INTRODUCTION platform and a wide pseudokeel or depressed keel at the aboral side of Pa elements. Traditionally species of the genus Siphonodella are used The shallow-water siphonodellids of the Chinese for biostratigraphy of the lower part of the Tournaisian branch appeared in the earliest Tournaisian and ranged (Sandberg et al. 1978; Ji 1985; Ji & Ziegler 1992; up to the late Tournaisian (Ji & Ziegler 1992). -
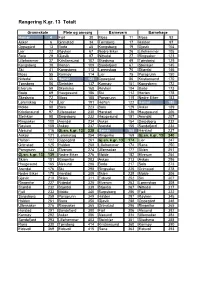
Rangering K.Gr. 13 Totalt
Rangering K.gr. 13 Totalt Grunnskole Pleie og omsorg Barnevern Barnehage Hamar 4 Fjell 30 Moss 11 Moss 92 Asker 6 Grimstad 34 Tønsberg 17 Halden 97 Oppegård 13 Bodø 45 Kongsberg 19 Gjøvik 104 Lier 22 Røyken 67 Nedre Eiker 26 Lillehammer 105 Sola 29 Gjøvik 97 Nittedal 27 Ringsaker 123 Lillehammer 37 Kristiansund 107 Skedsmo 49 Tønsberg 129 Kongsberg 38 Horten 109 Sandefjord 67 Steinkjer 145 Ski 41 Kongsberg 113 Lørenskog 70 Stjørdal 146 Moss 55 Karmøy 114 Lier 75 Porsgrunn 150 Nittedal 55 Hamar 123 Oppegård 86 Kristiansund 170 Tønsberg 56 Steinkjer 137 Karmøy 101 Kongsberg 172 Elverum 59 Skedsmo 168 Røyken 104 Bodø 173 Bodø 69 Haugesund 186 Ski 112 Horten 178 Skedsmo 72 Moss 188 Porsgrunn 115 Nedre Eiker 183 Lørenskog 74 Lier 191 Horten 122 Hamar 185 Molde 88 Sola 223 Sola 129 Asker 189 Kristiansund 97 Ullensaker 230 Harstad 136 Haugesund 206 Steinkjer 98 Sarpsborg 232 Haugesund 151 Arendal 207 Ringsaker 100 Arendal 234 Asker 154 Sarpsborg 232 Røyken 108 Askøy 237 Arendal 155 Sandefjord 234 Ålesund 116 Gj.sn. k.gr. 13 238 Hamar 168 Harstad 237 Askøy 121 Lørenskog 254 Ringerike 169 Gj.sn. k.gr. 13 240 Horten 122 Oppegård 261 Gj.sn. k.gr. 13 174 Lier 247 Grimstad 125 Halden 268 Lillehammer 174 Rana 250 Porsgrunn 133 Elverum 274 Ullensaker 177 Skien 251 Gj.sn. k.gr. 13 139 Nedre Eiker 276 Molde 182 Elverum 254 Skien 151 Ringerike 283 Askøy 213 Askøy 256 Haugesund 165 Ålesund 288 Bodø 217 Sola 273 Arendal 176 Ski 298 Ringsaker 225 Grimstad 278 Nedre Eiker 179 Harstad 309 Skien 239 Molde 306 Gjøvik 210 Skien 311 Eidsvoll 252 Ski 307 Ringerike -

Upper Ordovician Conodont Biostratigraphy and Revised Lithostratigraphy and Geological Map, Akpatok Island, Ungava Bay, Nunavut
Canadian Journal of Earth Sciences Upper Ordovician conodont biostratigraphy and revised lithostratigraphy and geological map, Akpatok Island, Ungava Bay, Nunavut Journal: Canadian Journal of Earth Sciences Manuscript ID cjes-2017-0145.R1 Manuscript Type: Article Date Submitted by the Author: 08-Sep-2017 Complete List of Authors: Zhang, Shunxin; Canada-Nunavut Geoscience Office, Is the invited manuscript for Draft consideration in a Special N/A Issue? : Upper Ordovician, conodont biostratigraphy, geological map, Akpatok Keyword: Island, Ungava Bay https://mc06.manuscriptcentral.com/cjes-pubs Page 1 of 55 Canadian Journal of Earth Sciences 1 2 Upper Ordovician conodont biostratigraphy and revised lithostratigraphy 3 and geological map, Akpatok Island, Ungava Bay, Nunavut 4 5 6 7 Shunxin Zhang 8 9 10 11 Canada - Nunavut Geoscience Office, PO Box 2319, 1106 Inuksugait IV, 1 st floor, Iqaluit, 12 Nunavut X0A 0H0, Canada; [email protected] 13 Draft 14 15 16 17 Correspondence author: 18 Shunxin Zhang 19 PO Box 2319, 1106 Inuksugait IV, 1 st floor, Iqaluit, Nunavut X0A 0H0, Canada; 20 Phone: (867) 975-4579 21 Fax: (867) 979-0708 22 Email: [email protected] 23 24 25 ESS contribution number: 20160440 26 1 https://mc06.manuscriptcentral.com/cjes-pubs Canadian Journal of Earth Sciences Page 2 of 55 27 Upper Ordovician conodont biostratigraphy and revised lithostratigraphy 28 and geological map, Akpatok Island, Ungava Bay, Nunavut 29 Shunxin Zhang 30 31 Abstract 32 Stratigraphic units exposed on Akpatok Island, Ungava Bay, Nunavut, were previously 33 recognized as Boas River and Akpatok formations; their biostratigraphic ages and correlations, 34 in particular the stratigraphic position and age of the organic rich “Boas River” Formation, were 35 largely based on limited data. -

Conodont Stratigraphy of a Highly Tectonised Silurian-Devonian Section in the San Basilio Area (SE Sardinia, Italy)
Bollettino della Societa Paleontologica Italiana 40 (3), 2001 ISSN 0375-7633 Modena, Dicembre 2001 Conodont stratigraphy of a highly tectonised Silurian-Devonian section in the San Basilio area (SE Sardinia, Italy) Carlo CORRADINI F rancesco LEONE Alfredo Loi Enrico SERPAGLI Dipartimento del Museo di Paleobiologia Dipartimento di Scienze della Terra Dipartimento del Museo di Paleobiologia e dell'Orto Botanico Universita di Cagliari e dell'Orto Botanico Universidt di Modena e Reggio Emilia Universita di Modena e Reggio Emilia KEYWORDS- Conodonts, Biostratigraphy, Tectonics, Silurian, Devonian, SE Sardinia. ABSTRACT- Several conodont species belonging to five "late Silurian biozones (Ancoradella ploeckensis, Polygnathoides siluricus, Ozarkodina crispa, Ozarkodina remscheidensis, Oulodus elegans detortus) and four Early and Late Devonian biozones (delta, pesavis, kitabicus-excavatus and Late rhenana) allow a better interpretation of stratigraphy and tectonics of the San Basilio area in western Gerrei. Frasnian sediments are documented for the first time in SE Sardinia. RIASSUNTO- [Stratigrafia a conodonti di una sezione forremente tettonizzata di eta Siluriano-Devoniano nell'area di San Basilio (Sardegna SE)] - Grazie alla biostratigrafia a conodonti viene interpretata la complessa tettonica di una sezione nell'area di San Basilio (Sardegna sud-orientale) e viene ricostruita la successione originaria. Sono documentate cinque biozone a conodonti del Siluriano superiore (Ancoradella eioeckensis, Polygnathoides siluricus, Ozarkodina crispa, -

Upcoming Projects Infrastructure Construction Division About Bane NOR Bane NOR Is a State-Owned Company Respon- Sible for the National Railway Infrastructure
1 Upcoming projects Infrastructure Construction Division About Bane NOR Bane NOR is a state-owned company respon- sible for the national railway infrastructure. Our mission is to ensure accessible railway infra- structure and efficient and user-friendly ser- vices, including the development of hubs and goods terminals. The company’s main responsible are: • Planning, development, administration, operation and maintenance of the national railway network • Traffic management • Administration and development of railway property Bane NOR has approximately 4,500 employees and the head office is based in Oslo, Norway. All plans and figures in this folder are preliminary and may be subject for change. 3 Never has more money been invested in Norwegian railway infrastructure. The InterCity rollout as described in this folder consists of several projects. These investments create great value for all travelers. In the coming years, departures will be more frequent, with reduced travel time within the InterCity operating area. We are living in an exciting and changing infrastructure environment, with a high activity level. Over the next three years Bane NOR plans to introduce contracts relating to a large number of mega projects to the market. Investment will continue until the InterCity rollout is completed as planned in 2034. Additionally, Bane NOR plans together with The Norwegian Public Roads Administration, to build a safer and faster rail and road system between Arna and Stanghelle on the Bergen Line (western part of Norway). We rely on close -

The Nature of Ordovician Limestone-Marl Alternations in the Oslo-Asker District
.re./sereprs OPEN The nature of Ordovician limestone-marl alternations in the Oslo-Asker District (Norway): reee: 1 e 01 epe: 0 eer 01 witnesses of primary glacio-eustasy se: 07 r 016 or diagenetic rhythms? Chloé E. A. Amberg1, Tim Collart2, Wout Salenbien2,3, Lisa M. Egger4,5, Axel Munnecke4, Arne T. Nielsen6, Claude Monnet1, Øyvind Hammer7 & Thijs R. A. Vandenbroucke1,2 Ordovician limestone-marl alternations in the Oslo-Asker District have been interpreted as signaling glacio-eustatic lowstands, which would support a prolonged “Early Palaeozoic Icehouse”. However, these rhythmites could alternatively refect diferential diagenesis, without sedimentary trigger. Here, we test both hypotheses through one Darriwilian and three Katian sections. Our methodology consists of a bed-by-bed analysis of palynological (chitinozoan) and geochemical (XRF) data, to evaluate whether the limestone/marl couplets refect an original cyclic signal. The results reveal similar palynomorph assemblages in limestones and marls. Exceptions, which could be interpreted as refecting palaeoclimatological fuctuations, exist at the species level: Ancyrochitina bornholmensis seems to be more abundant in the marl samples from the lower Frognerkilen Formation on Nakkholmen Island. However, these rare cases where chitinozoans difer between limestone/marl facies are deemed insufcient for the identifcation of original cyclicity. The geochemical data show a near-perfect correlation between insoluble elements in the limestone and the marls, which indicates a similar composition of the potential precursor sediment, also in the Frognerkilen Formation. This is consistent with the palynological data. Although an original cyclic pattern could still be recorded by other, uninvestigated parameters, our palaeontological and geochemical data combined do not support the presence of such a signal. -

Conodont Biostratigraphy of the Bakken and Lower Lodgepole Formations (Devonian and Mississippian), Williston Basin, North Dakota Timothy P
University of North Dakota UND Scholarly Commons Theses and Dissertations Theses, Dissertations, and Senior Projects 1986 Conodont biostratigraphy of the Bakken and lower Lodgepole Formations (Devonian and Mississippian), Williston Basin, North Dakota Timothy P. Huber University of North Dakota Follow this and additional works at: https://commons.und.edu/theses Part of the Geology Commons Recommended Citation Huber, Timothy P., "Conodont biostratigraphy of the Bakken and lower Lodgepole Formations (Devonian and Mississippian), Williston Basin, North Dakota" (1986). Theses and Dissertations. 145. https://commons.und.edu/theses/145 This Thesis is brought to you for free and open access by the Theses, Dissertations, and Senior Projects at UND Scholarly Commons. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of UND Scholarly Commons. For more information, please contact [email protected]. CONODONT BIOSTRATIGRAPHY OF THE BAKKEN AND LOWER LODGEPOLE FORMATIONS (DEVONIAN AND MISSISSIPPIAN), WILLISTON BASIN, NORTH DAKOTA by Timothy P, Huber Bachelor of Arts, University of Minnesota - Morris, 1983 A Thesis Submitted to the Graduate Faculty of the University of North Dakota in partial fulfillment of the requirements for the degree of Master of Science Grand Forks, North Dakota December 1986 This thesis submitted by Timothy P. Huber in partial fulfillment of the requirements for the Degree of Master of Science from the University of North Dakota has been read by the Faculty Advisory Committee under whom the work has been done, and is hereby approved. This thesis meets the standards for appearance and conforms to the style and format requirements of the Graduate School at the University of North Dakota and is hereby approved. -

Records of Carbon and Sulfur Cycling During the Silurian Ireviken Event in Gotland, Sweden
Accepted Manuscript Records of Carbon and Sulfur Cycling During the Silurian Ireviken Event in Gotland, Sweden Catherine V. Rose, Woodward W. Fischer, Seth Finnegan, David A. Fike PII: S0016-7037(18)30660-4 DOI: https://doi.org/10.1016/j.gca.2018.11.030 Reference: GCA 11025 To appear in: Geochimica et Cosmochimica Acta Received Date: 4 September 2018 Revised Date: 15 November 2018 Accepted Date: 21 November 2018 Please cite this article as: Rose, C.V., Fischer, W.W., Finnegan, S., Fike, D.A., Records of Carbon and Sulfur Cycling During the Silurian Ireviken Event in Gotland, Sweden, Geochimica et Cosmochimica Acta (2018), doi: https:// doi.org/10.1016/j.gca.2018.11.030 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. Records of Carbon and Sulfur Cycling During the Silurian Ireviken Event in Gotland, Sweden Catherine V. Rose1*, Woodward W. Fischer2, Seth Finnegan3, & David A. Fike4* 1School of Earth & Environmental Sciences, University of St Andrews, Fife, KY16 9AL, UK; 2Division of Geological & Planetary Sciences, California Institute of Technology, Pasadena, CA 91125 USA; 3Department of Integrative Biology, University of California, Berkeley, CA 94720-3140 USA; 4Department of Earth & Planetary Sciences, Washington University, St. -

(Mecoptera) in Norway
© Norwegian Journal of Entomology. 21 June 2011 Distribution of Boreus westwoodi Hagen, 1866 and Boreus hyemalis (L., 1767) (Mecoptera) in Norway SIGMUND HÅGVAR & EIVIND ØSTBYE Hågvar, S. & Østbye, E. 2011. Distribution of Boreus westwoodi Hagen, 1866 and Boreus hyemalis (L., 1767) (Mecoptera) in Norway. Norwegian Journal of Entomology 58, 73–80. An extensive material collected during nearly fifty years adds new detailed information on the distribution of the winter active insects Boreus westwoodi Hagen, 1866 and B. hyemalis (L., 1767) in Norway. Since females are difficult to identify, the new data rely on males. Based on the revised Strand-system, the following geographical regions are new to B. westwoodi: Ø, BØ, VAY, ON, TEI, TEY, MRI, MRY, and TRY. For B. hyemalis, AK, BØ, TEI, RY, SFI, and NTI are new regions. While B. westwoodi is widespread in Norway, including the three northernmost counties, B. hyemalis seems to be restricted to the south, with the northernmost record in NTI. In Sweden, the situation is similar: B. westwoodi is widespread, while B. hyemalis has been recorded as far north as Västerbotten, at a latitude corresponding to the northernmost record in Norway. The known distribution of both species in Norway is presented on EIS-grid map. Key words: Boreus hyemalis, Boreus westwoodi, Mecoptera, distribution, Norway. Sigmund Hågvar, Department of Ecology and Natural Resource Management, P.O. Box 5003, Norwegian University of Life Sciences, NO-1432 Ås, Norway. E-mail: [email protected] Eivind Østbye, Ringeriksveien 580, NO-3410 Sylling, Norway. E-mail: [email protected] Introduction county was described by Greve (1966).