Nuclear Fusion Enhances Cancer Cell Killing Efficacy in a Protontherapy Model
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Glossary Derived From: Human Research Program Integrated Research Plan, Revision A, (January 2009)
Glossary derived from: Human Research Program Integrated Research Plan, Revision A, (January 2009). National Aeronautics and Space Administration, Johnson Space Center, Houston, Texas 77058, pages 232-280. Report No. 153: Information Needed to Make Radiation Protection Recommendations for Space Missions Beyond Low-Earth Orbit (2006). National Council on Radiation Protection and Measurements, pages 309-318. Reprinted with permission of the National Council on Radiation Protection and Measurements, http://NCRPonline.org . Managing Space Radiation Risk in the New Era of Space Exploration (2008). Committee on the Evaluation of Radiation Shielding for Space Exploration, National Research Council. National Academies Press, pages 111-118. -A- AAPM: American Association of Physicists in Medicine. absolute risk: Expression of excess risk due to exposure as the arithmetic difference between the risk among those exposed and that obtaining in the absence of exposure. absorbed dose (D): Average amount of energy imparted by ionizing particles to a unit mass of irradiated material in a volume sufficiently small to disregard variations in the radiation field but sufficiently large to average over statistical fluctuations in energy deposition, and where energy imparted is the difference between energy entering the volume and energy leaving the volume. The same dose has different consequences depending on the type of radiation delivered. Unit: gray (Gy), equivalent to 1 J/kg. ACE: Advanced Composition Explorer Mission, launched in 1997 and orbiting the L1 libration point to sample energetic particles arriving from the Sun and interstellar and galactic sources. It also provides continuous coverage of solar wind parameters and solar energetic particle intensities (space weather). When reporting space weather, it can provide an advance warning (about one hour) of geomagnetic storms that can overload power grids, disrupt communications on Earth, and present a hazard to astronauts. -

Cherenkov Radiation
TheThe CherenkovCherenkov effecteffect A charged particle traveling in a dielectric medium with n>1 radiates Cherenkov radiation B Wave front if its velocity is larger than the C phase velocity of light v>c/n or > 1/n (threshold) A β Charged particle The emission is due to an asymmetric polarization of the medium in front and at the rear of the particle, giving rise to a varying electric dipole momentum. dN Some of the particle energy is convertedγ = 491into light. A coherent wave front is dx generated moving at velocity v at an angle Θc If the media is transparent the Cherenkov light can be detected. If the particle is ultra-relativistic β~1 Θc = const and has max value c t AB n 1 cosθc = = = In water Θc = 43˚, in ice 41AC˚ βct βn 37 TheThe CherenkovCherenkov effecteffect The intensity of the Cherenkov radiation (number of photons per unit length of particle path and per unit of wave length) 2 2 2 2 2 Number of photons/L and radiation d N 4π z e 1 2πz 2 = 2 1 − 2 2 = 2 α sin ΘC Wavelength depends on charge dxdλ hcλ n β λ and velocity of particle 2πe2 α = Since the intensity is proportional to hc 1/λ2 short wavelengths dominate dN Using light detectors (photomultipliers)γ = sensitive491 in 400-700 nm for an ideally 100% efficient detector in the visibledx € 2 dNγ λ2 d Nγ 2 2 λ2 dλ 2 2 11 1 22 2 d 2 z sin 2 z sin 490393 zz sinsinΘc photons / cm = ∫ λ = π α ΘC ∫ 2 = π α ΘC 2 −− 2 = α ΘC λ1 λ1 dx dxdλ λ λλ1 λ2 d 2 N d 2 N dλ λ2 d 2 N = = dxdE dxdλ dE 2πhc dxdλ Energy loss is about 104 less hc 2πhc than 2 MeV/cm in water from € -

Digital Physics: Science, Technology and Applications
Prof. Kim Molvig April 20, 2006: 22.012 Fusion Seminar (MIT) DDD-T--TT FusionFusion D +T → α + n +17.6 MeV 3.5MeV 14.1MeV • What is GOOD about this reaction? – Highest specific energy of ALL nuclear reactions – Lowest temperature for sizeable reaction rate • What is BAD about this reaction? – NEUTRONS => activation of confining vessel and resultant radioactivity – Neutron energy must be thermally converted (inefficiently) to electricity – Deuterium must be separated from seawater – Tritium must be bred April 20, 2006: 22.012 Fusion Seminar (MIT) ConsiderConsider AnotherAnother NuclearNuclear ReactionReaction p+11B → 3α + 8.7 MeV • What is GOOD about this reaction? – Aneutronic (No neutrons => no radioactivity!) – Direct electrical conversion of output energy (reactants all charged particles) – Fuels ubiquitous in nature • What is BAD about this reaction? – High Temperatures required (why?) – Difficulty of confinement (technology immature relative to Tokamaks) April 20, 2006: 22.012 Fusion Seminar (MIT) DTDT FusionFusion –– VisualVisualVisual PicturePicture Figure by MIT OCW. April 20, 2006: 22.012 Fusion Seminar (MIT) EnergeticsEnergetics ofofof FusionFusion e2 V ≅ ≅ 400 KeV Coul R + R V D T QM “tunneling” required . Ekin r Empirical fit to data 2 −VNuc ≅ −50 MeV −2 A1 = 45.95, A2 = 50200, A3 =1.368×10 , A4 =1.076, A5 = 409 Coefficients for DT (E in KeV, σ in barns) April 20, 2006: 22.012 Fusion Seminar (MIT) TunnelingTunneling FusionFusion CrossCross SectionSection andand ReactivityReactivity Gamow factor . Compare to DT . April 20, 2006: 22.012 Fusion Seminar (MIT) ReactivityReactivity forfor DTDT FuelFuel 8 ] 6 c e s / 3 m c 6 1 - 0 4 1 x [ ) ν σ ( 2 0 0 50 100 150 200 T1 (KeV) April 20, 2006: 22.012 Fusion Seminar (MIT) Figure by MIT OCW. -

Radiation and Risk: Expert Perspectives Radiation and Risk: Expert Perspectives SP001-1
Radiation and Risk: Expert Perspectives Radiation and Risk: Expert Perspectives SP001-1 Published by Health Physics Society 1313 Dolley Madison Blvd. Suite 402 McLean, VA 22101 Disclaimer Statements and opinions expressed in publications of the Health Physics Society or in presentations given during its regular meetings are those of the author(s) and do not necessarily reflect the official position of the Health Physics Society, the editors, or the organizations with which the authors are affiliated. The editor(s), publisher, and Society disclaim any responsibility or liability for such material and do not guarantee, warrant, or endorse any product or service mentioned. Official positions of the Society are established only by its Board of Directors. Copyright © 2017 by the Health Physics Society All rights reserved. No part of this publication may be reproduced or distributed in any form, in an electronic retrieval system or otherwise, without prior written permission of the publisher. Printed in the United States of America SP001-1, revised 2017 Radiation and Risk: Expert Perspectives Table of Contents Foreword……………………………………………………………………………………………………………... 2 A Primer on Ionizing Radiation……………………………………………………………………………... 6 Growing Importance of Nuclear Technology in Medicine……………………………………….. 16 Distinguishing Risk: Use and Overuse of Radiation in Medicine………………………………. 22 Nuclear Energy: The Environmental Context…………………………………………………………. 27 Nuclear Power in the United States: Safety, Emergency Response Planning, and Continuous Learning…………………………………………………………………………………………….. 33 Radiation Risk: Used Nuclear Fuel and Radioactive Waste Disposal………………………... 42 Radiation Risk: Communicating to the Public………………………………………………………… 45 After Fukushima: Implications for Public Policy and Communications……………………. 51 Appendix 1: Radiation Units and Measurements……………………………………………………. 57 Appendix 2: Half-Life of Some Radionuclides…………………………………………………………. 58 Bernard L. -
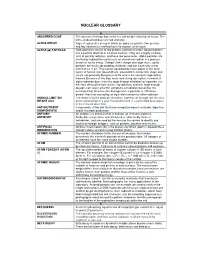
Nuclear Glossary
NUCLEAR GLOSSARY A ABSORBED DOSE The amount of energy deposited in a unit weight of biological tissue. The units of absorbed dose are rad and gray. ALPHA DECAY Type of radioactive decay in which an alpha ( α) particle (two protons and two neutrons) is emitted from the nucleus of an atom. ALPHA (ααα) PARTICLE. Alpha particles consist of two protons and two neutrons bound together into a particle identical to a helium nucleus. They are a highly ionizing form of particle radiation, and have low penetration. Alpha particles are emitted by radioactive nuclei such as uranium or radium in a process known as alpha decay. Owing to their charge and large mass, alpha particles are easily absorbed by materials and can travel only a few centimetres in air. They can be absorbed by tissue paper or the outer layers of human skin (about 40 µm, equivalent to a few cells deep) and so are not generally dangerous to life unless the source is ingested or inhaled. Because of this high mass and strong absorption, however, if alpha radiation does enter the body through inhalation or ingestion, it is the most destructive form of ionizing radiation, and with large enough dosage, can cause all of the symptoms of radiation poisoning. It is estimated that chromosome damage from α particles is 100 times greater than that caused by an equivalent amount of other radiation. ANNUAL LIMIT ON The intake in to the body by inhalation, ingestion or through the skin of a INTAKE (ALI) given radionuclide in a year that would result in a committed dose equal to the relevant dose limit . -

Radiation and Your Patient: a Guide for Medical Practitioners
RADIATION AND YOUR PATIENT: A GUIDE FOR MEDICAL PRACTITIONERS A web module produced by Committee 3 of the International Commission on Radiological Protection (ICRP) What is the purpose of this document ? In the past 100 years, diagnostic radiology, nuclear medicine and radiation therapy have evolved from the original crude practices to advanced techniques that form an essential tool for all branches and specialties of medicine. The inherent properties of ionising radiation provide many benefits but also may cause potential harm. In the practice of medicine, there must be a judgement made concerning the benefit/risk ratio. This requires not only knowledge of medicine but also of the radiation risks. This document is designed to provide basic information on radiation mechanisms, the dose from various medical radiation sources, the magnitude and type of risk, as well as answers to commonly asked questions (e.g radiation and pregnancy). As a matter of ease in reading, the text is in a question and answer format. Interventional cardiologists, radiologists, orthopaedic and vascular surgeons and others, who actually operate medical x-ray equipment or use radiation sources, should possess more information on proper technique and dose management than is contained here. However, this text may provide a useful starting point. The most common ionising radiations used in medicine are X, gamma, beta rays and electrons. Ionising radiation is only one part of the electromagnetic spectrum. There are numerous other radiations (e.g. visible light, infrared waves, high frequency and radiofrequency electromagnetic waves) that do not posses the ability to ionize atoms of the absorbing matter. -

Literature Survey on Decorporation of Radionuclides from the Human Body
Literature Survey on Decorporation of Radionuclides from the Human Body E.A. Waller, R.Z. Stodilka, K. Leach and L. Prud’homme-Lalonde Defence R&D Canada - Ottawa TECHNICAL MEMORANDUM DRDC Ottawa TM 2002-042 April 2002 Literature Survey on Decorporation of Radionuclides from the Human Body E.A. Waller SAIC Canada, Inc R.Z. Stodilka, K. Leach and L. Prud’homme-Lalonde Space Systems and Technology Defence R&D Canada - Ottawa Technical Memorandum DRDC Ottawa TM 2002-042 April 2002 © Her Majesty the Queen as represented by the Minister of National Defence, 2002 © Sa majesté la reine, représentée par le ministre de la Défense nationale, 2002 Abstract The broad use of radionuclides by many industries has greatly increased the probability of events that could lead to internalized contamination. Examples include accidents and/or intentional damage to nuclear power plants or radiation therapy units in hospitals, the use of radiological dispersal weapons, and lost or stolen radionuclide sources. Developing effective countermeasures requires knowledge of the physical and chemical composition of the radionuclides, their metabolic activities within the body, and methods to expedite their elimination from the body. This report presents a summary of information pertaining to intake and decorporation of radionuclides from humans. This information would be the first step in establishing a field protocol to guide physicians in military missions. Developing such a guide requires an understanding of the dangers associated with internal radioisotope contamination, decision levels for administering therapy (risk vs. benefit) and protocols for administering therapy. As presented, this study could be used to decide what decorporation pharmaceuticals should be maintained in quantity by the military, and how to best train officers with medical responsibilities. -

And Stereotactic Body Radiation Therapy (SBRT)
Geisinger Health Plan Policies and Procedure Manual Policy: MP084 Section: Medical Benefit Policy Subject: Stereotactic Radiosurgery and Stereotactic Body Radiation Therapy I. Policy: Stereotactic Radiosurgery (SRS) and Stereotactic Body Radiation Therapy (SBRT) II. Purpose/Objective: To provide a policy of coverage regarding Stereotactic Radiosurgery (SRS) and Stereotactic Body Radiation Therapy (SBRT) III. Responsibility: A. Medical Directors B. Medical Management IV. Required Definitions 1. Attachment – a supporting document that is developed and maintained by the policy writer or department requiring/authoring the policy. 2. Exhibit – a supporting document developed and maintained in a department other than the department requiring/authoring the policy. 3. Devised – the date the policy was implemented. 4. Revised – the date of every revision to the policy, including typographical and grammatical changes. 5. Reviewed – the date documenting the annual review if the policy has no revisions necessary. V. Additional Definitions Medical Necessity or Medically Necessary means Covered Services rendered by a Health Care Provider that the Plan determines are: a. appropriate for the symptoms and diagnosis or treatment of the Member's condition, illness, disease or injury; b. provided for the diagnosis, and the direct care and treatment of the Member's condition, illness disease or injury; c. in accordance with current standards of good medical treatment practiced by the general medical community. d. not primarily for the convenience of the Member, or the Member's Health Care Provider; and e. the most appropriate source or level of service that can safely be provided to the Member. When applied to hospitalization, this further means that the Member requires acute care as an inpatient due to the nature of the services rendered or the Member's condition, and the Member cannot receive safe or adequate care as an outpatient. -

New Discoveries in Radiation Science
cancers Editorial New Discoveries in Radiation Science Géza Sáfrány 1,*, Katalin Lumniczky 1 and Lorenzo Manti 2 1 Department Radiobiology and Radiohygiene, National Public Health Center, 1221 Budapest, Hungary; [email protected] 2 Department of Physics, University of Naples Federico II, 80126 Naples, Italy; [email protected] * Correspondence: [email protected]; Tel.: +36-309199218 This series of 16 articles (8 original articles and 8 reviews) was written by internation- ally recognized scientists attending the 44th Congress of the European Radiation Research Society (Pécs, Hungary). Ionizing radiation is an interesting agent because it is used to cure cancers and can also induce cancer. The effects of ionizing radiation at the organism level depend on the response of the cells. When radiation hits a cell, it might damage any cellular organelles and macromolecules. Unrepairable damage leads to cell death, while misrepaired alterations leave mutations in surviving cells. If the repair is errorless, normal cells will survive. However, in a small percentage of the seemingly healthy cells the number of spontaneous mutations will increase, which is a sign of radiation-induced genomic instability. Radiation-induced cell death is behind the development of acute radiation syndromes and the killing of tumorous and normal cells during radiation therapy. Radiation-induced mutations in surviving cells might lead to the induction of tumors. According to the central paradigm of radiation biology, the genetic material, that is the DNA, is the main cellular target of ionizing radiation. Many different types of damage are induced by radiation in the DNA, but the most deleterious effects arise from double strand breaks (DSBs). -

Radiation Therapy
Radiation Therapy What is radiation therapy? Radiation therapy is the use of high-energy x-rays or other particles to destroy cancer cells. A doctor who uses radiation therapy to treat cancer is called a radiation oncologist. The goal of radiation therapy is to destroy the cancer cells without harming nearby healthy tissue. It may be used along with other cancer treatments or as the main treatment. Sometimes radiation therapy is used to relieve symptoms, called palliative radiation therapy. More than half of all people with cancer receive some type of radiation therapy. What are the different types of radiation therapy? The most common type is called external-beam radiation therapy, which is radiation given from a machine located outside the body. Types of external-beam radiation therapy include proton therapy, 3-dimensional conformal radiation therapy (3D-CRT), intensity-modulated radiation therapy (IMRT), and stereotactic radiation therapy. Sometimes radiation therapy involves bringing a radioactive source close to a tumor. This is called internal radiation therapy or brachytherapy. The type you receive depends on many factors. Learn more about radiation treatment at www.cancer.net/radiationtherapy. What should I expect during radiation therapy? Before treatment begins, you will meet with the radiation oncologist to review your medical history and discuss the potential risks and benefits. If you choose to receive radiation therapy, you may undergo tests to plan the treatment and evaluate the results. Before radiation therapy begins, it must be planned carefully. This planning stage is often called a “simulation.” During this visit, the medical team will figure out the best position for you to be in during treatment. -

Maximizing Specific Energy by Breeding Deuterium
Maximizing specific energy by breeding deuterium Justin Ball Ecole Polytechnique F´ed´eralede Lausanne (EPFL), Swiss Plasma Center (SPC), CH-1015 Lausanne, Switzerland E-mail: [email protected] Abstract. Specific energy (i.e. energy per unit mass) is one of the most fundamental and consequential properties of a fuel source. In this work, a systematic study of measured fusion cross-sections is performed to determine which reactions are potentially feasible and identify the fuel cycle that maximizes specific energy. This reveals that, by using normal hydrogen to breed deuterium via neutron capture, the conventional catalyzed D-D fusion fuel cycle can attain a specific energy greater than anything else. Simply surrounding a catalyzed D-D reactor with water enables deuterium fuel, the dominant stockpile of energy on Earth, to produce as much as 65% more energy. Lastly, the impact on space propulsion is considered, revealing that an effective exhaust velocity exceeding that of deuterium-helium-3 is theoretically possible. 1. Introduction The aim of this work is to investigate the basic question \What fuel source can provide the most energy with the least mass?" For this question, we will look into the near future and assume that the current incremental and steady development of technology will continue. However, we will not postulate any breakthroughs or new physical processes because they cannot be predicted with any certainty. It is already well-known that the answer to our question must involve fusion fuels [1,2], but nothing more precise has been rigorously establishedz. A more precise answer may be useful in the future arXiv:1908.00834v1 [physics.pop-ph] 2 Aug 2019 when the technology required to implement it becomes available. -

Exposure Data
X-RADIATION AND γ-RADIATION 1. Exposure data 1.1 Occurrence 1.1.1 X-radiation X-rays are electromagnetic waves in the spectral range between the shortest ultraviolet (down to a few tens of electron volts) and γ-radiation (up to a few tens of mega electron volts) (see Figure 2, Overall introduction). The term γ-radiation is usually restricted to radiation originating from the atomic nucleus and from particle annihilation, while the term X-radiation covers photon emissions from electron shells. X-rays are emitted when charged particles are accelerated or decelerated, during transitions of electrons from the outer regions of the atomic shell to regions closer to the nucleus, and as bremsstrahlung, i.e. radiation produced when an electron collides with, or is deflected by, a positively charged nucleus. The resulting line spectra are characteristic for the corresponding element, whereas bremsstrahlung shows a conti- nuous spectrum with a steep border at the shortest wavelengths. Interaction of X-rays with matter is described by the Compton scattering and photoelectric effect and their resulting ionizing potentials, which lead to significant chemical and biological effects. Ions and radicals are produced in tissues from single photons and cause degradation and changes in covalent binding in macromolecules such as DNA. In other parts of the electromagnetic spectrum, below the spectra of ultraviolet and visible light, the single photon energies are too low to cause genotoxic × –μ⋅d effects. The intensity (I) of X-rays inside matter decreases according to I = I0 10 , where d is the depth and μ a coefficient specific to the interacting material and the corresponding wavelength.