Lecture Notebook
to accompany
- Sinauer aSSociateS, inc.
- MacMillan
Copyright © 2014 Sinauer Associates, Inc. Cover photograph © Alex Mustard/naturepl.com. This document may not be modified or distributed (either electronically or on paper) without the permission of the publisher, with the following exception: Individual users may enter their own notes into this document and may print it for their own personal use.
The Origin and Diversification of Eukaryotes
0207
27.1 A Hypothetical Sequence for the Evolution of the Eukaryotic Cell (Page 550)
Cell wall DNA
The protective cell wall was lost.
1
Infolding of the
2
plasma membrane added surface area without increasing the cell’s volume.
Cytoskeleton (microfilament and microtubules) formed.
34
Internal membranes studded with ribosomes formed.
As regions of the infolded plasma membrane enclosed the cell’s DNA, a precursor of a
5
nucleus formed.
Microtubules from the cytoskeleton formed the eukaryotic flagellum, enabling propulsion.
6
Early digestive vacuoles evolved into lysosomes using enzymes from the early endoplasmic reticulum.
789
Mitochondria formed through endosymbiosis with a proteobacterium.
Endosymbiosis with cyanobacteria led to the development of chloroplasts.
Flagellum
To add your own notes to any page, use Adobe Reader’s
Typewriter feature, accessible via the Typewriter bar at the top of the window. (Requires Adobe Reader 8 or later. Adobe Reader can be downloaded free of charge from the
Adobe website: http://get.adobe.com/reader.)
Chloroplast Mitochondrion
Nucleus
2
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
3
(A) Primary endosymbiosis
Eukaryote
Cyanobacterium
Cyanobacterium outer membrane
Peptidoglycan Cyanobacterium inner membrane
Host cell nucleus
Chloroplast
Peptidoglycan has been lost except in glaucophytes.
Chloroplastcontaining eukaryotic cell
(B) Secondary endosymbiosis
Host eukaryotic cell
Host membrane (from endocytosis) encloses the engulfed cell.
A trace of the engulfed cell’s nucleus is retained in some groups.
The engulfed cell’s plasma membrane (white) has been lost in euglenids and dinoflagellates.
27.2 Endosymbiotic Events in the Evolution of Chloroplasts
(Page 552)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
4
Alveolates
Stramenopiles
Rhizaria Excavates Plantae Chs. 28 and 29
Amoebozoans
Fungi
Ch. 30
Ophisthokonts
Choanoflagellates
Animals
Chs. 31–33
Precambrian
- Paleozoic
- Mesozoic
Cenozoic
to 4.5 bya
- 1.5
- 1.4
- 1.3
- 1.2
- 1.1
- 1.0
- 0.9
- 0.8
- 0.7
- 0.6
- 0.5
- 0.4
- 0.3
- 0.2
- 0.1
- 0
Billions of years ago
27.3 Precambrian Divergence of Major Eukaryote Groups (Page 553)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
5
Dinoflagellates Apicomplexans
Ciliates
Stramenopiles
Rhizaria
In-Text Art (Page 553)
Peridinium sp.
Equatorial groove
Longitudinal groove
27.4 A Dinoflagellate (Page 554)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
6
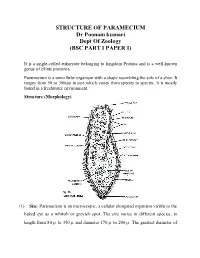
- (A) Paramecium sp.
- (B) Didinium nasutum
- (C) Euplotes sp.
10 µm
27.5 Diversity among the Ciliates (Page 554)
- 7 µm
- 25 µm
- Rows of fused cilia
- Oral groove
- Cilia
- Bands of cilia
The macronucleus controls the cell’s activities.
Micronuclei function in genetic recombination.
Contractile vacuole
Alveoli
Cilia
Digestive vacuole
Oral groove
Trichocyst Fibrils
Anal pore
Pellicle
Alveolus
Cilium
27.6 Anatomy of Paramecium (Page 555)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
7
INVESTIGATINGLIFE
27.7 The Role of Vacuoles in Ciliate Digestion
HYPOTHESIS The digestive vacuoles of Paramecium produce an acidic environment that allows the organism to digest food particles.
Method 1. Feed Paramecium yeast cells stained with Congo red, a dye that is red at neutral or basic pH but turns green at acidic pH.
2. Under a light microscope, observe the formation and degradation of digestive vacuoles within the Paramecium. Note time and sequence of color (i.e., acid level) changes.
Results
1 A digestive vacuole forms around yeast cells.
2 The change in color shows that the interior vacuole has become
Stained yeast cells
acidic.
Oral groove
As products of digestion move into the cytosol, the pH increases in the vacuole (the dye becomes red again).
3
Red-stained (basic) waste material is expelled.
4
CONCLUSION Some ciliates acidify digestive vacuoles to assist in the breakdown of food.
Go to BioPortal for discussion and relevant links for all
INVESTIGATINGLIFE figures.
(Page 555)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
8
Alveolates
Brown algae
Diatoms Oomycetes
Rhizaria
In-Text Art (Page 555)
25 µm
Diatoms display either radial (circular) symmetry...
...or bilateral (left-right) symmetry.
27.8 Diatom Diversity (Page 556)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
9
- (A) Himanthalia elongata
- (B) Postelsia palmiformis
Holdfasts
27.9 Brown Algae (Page 556)
Saprolegnia sp.
3 mm
27.10 An Oomycete (Page 557)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
10
Alveolates
Stramenopiles
Cercozoans Foraminiferans Radiolarians
In-Text Art (Page 557)
1 mm
27.11 Building Blocks of Limestone (Page 557)
- (A)
- (B)
- Astrolithium sp.
- Hexacontium sp.
50 µm
250 µm
27.12 Radiolarians Exhibit Distinctive Pseudopods and Radial Symmetry (Page 557)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
11
Excavates
Diplomonads Parabasalids
Heteroloboseans
Euglenids
Kinetoplastids
In-Text Art (Page 558)
(A) Giardia sp.
2.5 µm
(B) T r ichomonas vaginalis
2.5 µm
27.13 Some Excavate Groups Lack Mitochondria (Page 558)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
12
Photosynthetic chloroplasts are prominent features in a typical Euglena cell.
Flagella
Nucleus
Pigment shield
Stored polysaccharides from photosynthesis
Contractile vacuole
Photoreceptor
27.14 A Photosynthetic Euglenid (Page 559)
TablE27.1
Three Pathogenic Trypanosomes
- Trypanosoma brucei
- Trypanosoma cruzi
- Leishmania major
Leishmaniasis Sand fly
- Human disease
- Sleeping sickness
Tsetse fly
Chagas disease Assassin bugs (many species) None
Insect vector Vaccine or effective cure Strategy for survival
- None
- None
Changes surface recognition Causes changes in surface recognition Reduces effectiveness of macrophage
- molecules frequently
- molecules on host cell
- hosts
- Site in human body
- Bloodstream; in final stages, Enters cells, especially muscle cells
attacks nerve tissue
Enters cells, primarily macrophages
Approximate number of deaths per year
- 50,000
- 45,000
- 60,000
(Page 559)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
13
Amoebozoans
Loboseans
Plasmodial slime molds Cellular slime molds
In-Text Art (Page 559)
Chaos carolinensis
Pseudopods
Nebela collaris
120 µm
27.15 An Amoeba in Motion (Page 559)
Shell (test) made of sand grains
Plasma membrane of amoeba
Pseudopods of amoeba
18 µm
27.16 Life in a Glass House (Page 560)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
14
(A)
30 mm
(B)
1.5 mm
27.17 A Plasmodial Slime Mold
(Page 560)
The sporangium of the mature fruiting structure will release spores.
Dictyostelium discoideum
Fruiting structure (various stages)
Slug
0.25 mm
27.18 A Cellular Slime Mold (Page 561)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
15
Macronucleus Micronucleus
1 Two paramecia conjugate; all but one micronucleus in each cell disintegrate. The remaining micronucleus undergoes meiosis.
2 Three of the four haploid micronuclei disintegrate; the remaining micronucleus undergoes mitosis.
3
The paramecia donate micronuclei to each other. The macronuclei disintegrate.
4
The two micronuclei in each cell—each genetically
5 The new diploid micronuclei divide mitotically, eventually giving rise to a macronucleus and the appropriate number of micronuclei. different—fuse.
27.19 Conjugation in Paramecia (Page 562)
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
16
START
Eventually, some merozoites develop into male and
8
A blood-feeding female mosquito ingests the
Plasmodium gametocytes.
1
Merozoites also invade red blood cells, grow and divide, and lyse the cells. They can reinfect the liver, producing new generations.
7
female gametocytes.
Male gamete
Within the mosquito, male and female
gametocytes develop
into gametes, which fuse.
2
Red blood cell
Female gamete
The resulting zygote
3
enters the mosquito’s gut wall and forms a cyst.
- Events in human
- Events in mosquito
Mosquito's gut wall
The zygote gives rise to sporozoites that invade the salivary gland.
4
Sporozoites penetrate liver cells and develop into merozoites.
6
(B)
Human liver cell
Mosquito's salivary gland
The mosquito injects sporozoites into a human’s blood when it feeds.
5
Cysts
27.20 Life Cycle of the Malarial Parasite (Page 564)
Mosquito's gut wall
170 µm
© 2014 Sinauer Associates, Inc.
|
chapter 27 the origin and Diversification of eukaryotes
17
INVESTIGATINGLIFE
27.21 Can Corals Reacquire Dinoflagellate Endosymbionts Lost to Bleaching?
HYPOTHESIS Bleached corals can acquire new photosynthetic endosymbionts from their environment.
Method 1. Count numbers of Symbiodinium, a photosynthetic dinoflagellate, living symbiotically in samples of a coral (Briareum sp.).
2. Stimulate bleaching by maintaining all Briareum colonies in darkness for 12 weeks.
3. After 12 weeks of darkness, count numbers of Symbiodinium in the coral samples; then return all colonies to light.
4. In some of the bleached colonies (the experimental group), introduce Symbiodinium strain B211—dinoflagellates that contain a unique molecular marker. Do not expose the others (the control group) to strain B211. Maintain both groups in the light for 6 weeks.
Results
70
Experimental (exposed to strain B211)
60
Control (not exposed to strain B211)
50 40 30 20 10
Six weeks after return to light, both groups showed increases in number of symbionts present. DNA analysis showed that strain B211 symbionts were present in the experimental group.
After 12 weeks in dark, 0–1% of the photosynthetic endosymbionts remained.
3210
- Pre-bleach
- Post-bleach
- Week 3
- Week 6
(original state)
- Pre-bleach
- Post-bleach
CONCLUSION Corals can acquire new endosymbionts from their environment following bleaching.
Go to BioPortal for discussion and relevant links for all
INVESTIGATINGLIFE figures.
(Page 565)
© 2014 Sinauer Associates, Inc.