An Immunohistochemical Study of Lymphatic Elements in the Human Brain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nerve Ultrasound in Dorsal Root Ganglion Disorders: Smaller Nerves Lead to Bigger Insights
Clinical Neurophysiology 130 (2019) 550–551 Contents lists available at ScienceDirect Clinical Neurophysiology journal homepage: www.elsevier.com/locate/clinph Editorial Nerve ultrasound in dorsal root ganglion disorders: Smaller nerves lead to bigger insights See Article, pages 568–572 After decades of having to make do with electric stimulation representing the fascicles, bundled together in a large outer cable and recording (i.e. nerve conduction studies, electromyography sheath (van Alfen et al., 2018). and evoked potentials), nerve ultrasound now provides the oppor- Next, it is important to realize what the ratio between axon/ tunity to improve neurodiagnostic patient care by deploying a myelin and connective tissue in a given nerve segment is, and powerful tool to detect neuromuscular pathology in an accurate how that ratio changes from the proximal root to the distal end and patient-friendly way (Mah et al., 2018; Walker et al., 2018). branches (Schraut et al., 2016). Connective tissue elements of the Nerve ultrasound is also increasingly providing neurologists and perineurium and epineurium are relatively sparse at the very prox- clinical neurophysiologists with the opportunity to increase their imal root and plexus levels, with an average connective tissue con- insight in the pathophysiology of peripheral nervous system tent of around 25–30%. Ultrasonographically, this means that roots (PNS) pathology. In this issue of Clinical Neurophysiology, Leadbet- will always look rather black in appearance without much dis- ter and coworkers (Leadbetter et al., 2019) describe the results of cernible fascicular architecture, as the sparseness of connective tis- their study on nerve ultrasound for diagnosing sensory neuronopa- sue elements provides relatively few reflectors to create an image thy in spinocerebellar ataxia type 2 and CANVAS syndrome. -

Distance Learning Program Anatomy of the Human Brain/Sheep Brain Dissection
Distance Learning Program Anatomy of the Human Brain/Sheep Brain Dissection This guide is for middle and high school students participating in AIMS Anatomy of the Human Brain and Sheep Brain Dissections. Programs will be presented by an AIMS Anatomy Specialist. In this activity students will become more familiar with the anatomical structures of the human brain by observing, studying, and examining human specimens. The primary focus is on the anatomy, function, and pathology. Those students participating in Sheep Brain Dissections will have the opportunity to dissect and compare anatomical structures. At the end of this document, you will find anatomical diagrams, vocabulary review, and pre/post tests for your students. The following topics will be covered: 1. The neurons and supporting cells of the nervous system 2. Organization of the nervous system (the central and peripheral nervous systems) 4. Protective coverings of the brain 5. Brain Anatomy, including cerebral hemispheres, cerebellum and brain stem 6. Spinal Cord Anatomy 7. Cranial and spinal nerves Objectives: The student will be able to: 1. Define the selected terms associated with the human brain and spinal cord; 2. Identify the protective structures of the brain; 3. Identify the four lobes of the brain; 4. Explain the correlation between brain surface area, structure and brain function. 5. Discuss common neurological disorders and treatments. 6. Describe the effects of drug and alcohol on the brain. 7. Correctly label a diagram of the human brain National Science Education -

Review Article Meninges: from Protective Membrane to Stem Cell Niche
Am J Stem Cell 2012;1(2):92-105 www.AJSC.us /ISSN: 2160-4150/AJSC1205003 Review Article Meninges: from protective membrane to stem cell niche Ilaria Decimo1, Guido Fumagalli1, Valeria Berton1, Mauro Krampera2, Francesco Bifari2 1Department of Public Health and Community Medicine, Section of Pharmacology, University of Verona, Italy; 2De- partment of Medicine, Stem Cell Laboratory, Section of Hematology, University of Verona, Italy Received May 16, 2012; accepted May 23, 2012; Epub 28, 2012; Published June 30, 2012 Abstract: Meninges are a three tissue membrane primarily known as coverings of the brain. More in depth studies on meningeal function and ultrastructure have recently changed the view of meninges as a merely protective mem- brane. Accurate evaluation of the anatomical distribution in the CNS reveals that meninges largely penetrate inside the neural tissue. Meninges enter the CNS by projecting between structures, in the stroma of choroid plexus and form the perivascular space (Virchow-Robin) of every parenchymal vessel. Thus, meninges may modulate most of the physiological and pathological events of the CNS throughout the life. Meninges are present since the very early em- bryonic stages of cortical development and appear to be necessary for normal corticogenesis and brain structures formation. In adulthood meninges contribute to neural tissue homeostasis by secreting several trophic factors includ- ing FGF2 and SDF-1. Recently, for the first time, we have identified the presence of a stem cell population with neural differentiation potential in meninges. In addition, we and other groups have further described the presence in men- inges of injury responsive neural precursors. In this review we will give a comprehensive view of meninges and their multiple roles in the context of a functional network with the neural tissue. -

Endoscopic Anatomical Study of the Arachnoid Architecture on the Base of the Skull
DOI 10.1515/ins-2012-0005 Innovative Neurosurgery 2013; 1(1): 55–66 Original Research Article Peter Kurucz* , Gabor Baksa , Lajos Patonay and Nikolai J. Hopf Endoscopic anatomical study of the arachnoid architecture on the base of the skull. Part I: The anterior and middle cranial fossa Abstract: Minimally invasive neurosurgery requires a Introduction detailed knowledge of microstructures, such as the arach- noid membranes. In spite of many articles addressing The arachnoid was discovered and named by Gerardus arachnoid membranes, its detailed organization is still not Blasius in 1664 [ 22 ]. Key and Retzius were the first who well described. The aim of this study is to investigate the studied its detailed anatomy in 1875 [ 11 ]. This description was topography of the arachnoid in the anterior cranial fossa an anatomical one, without mentioning clinical aspects. The and the middle cranial fossa. Rigid endoscopes were intro- first clinically relevant study was provided by Liliequist in duced through defined keyhole craniotomies, to explore 1959 [ 13 ]. He described the radiological anatomy of the sub- the arachnoid structures in 110 fresh human cadavers. We arachnoid cisterns and mentioned a curtain-like membrane describe the topography and relationship to neurovascu- between the supra- and infratentorial cranial space bearing lar structures and suggest an intuitive terminology of the his name still today. Lang gave a similar description of the arachnoid. We demonstrate an “ arachnoid membrane sys- subarachnoid cisterns in 1973 [ 12 ]. With the introduction of tem ” , which consists of the outer arachnoid and 23 inner microtechniques in neurosurgery, the detailed knowledge arachnoid membranes in the anterior fossa and the middle of the surgical anatomy of the cisterns became more impor- fossa. -

The Strain Rates in the Brain, Brainstem, Dura, and Skull Under Dynamic Loadings
Mathematical and Computational Applications Article The Strain Rates in the Brain, Brainstem, Dura, and Skull under Dynamic Loadings Mohammad Hosseini-Farid 1,2,* , MaryamSadat Amiri-Tehrani-Zadeh 3, Mohammadreza Ramzanpour 1, Mariusz Ziejewski 1 and Ghodrat Karami 1 1 Department of Mechanical Engineering, North Dakota State University, Fargo, ND 58104, USA; [email protected] (M.R.); [email protected] (M.Z.); [email protected] (G.K.) 2 Department of Orthopedic Surgery, Mayo Clinic, Rochester, MN 55905, USA 3 Department of Computer Science, North Dakota State University, Fargo, ND 58104, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-7012315859 Received: 7 March 2020; Accepted: 5 April 2020; Published: 7 April 2020 Abstract: Knowing the precise material properties of intracranial head organs is crucial for studying the biomechanics of head injury. It has been shown that these biological tissues are significantly rate-dependent; hence, their material properties should be determined with respect to the range of deformation rate they experience. In this paper, a validated finite element human head model is used to investigate the biomechanics of the head in impact and blast, leading to traumatic brain injuries (TBI). We simulate the head under various directions and velocities of impacts, as well as helmeted and unhelmeted head under blast shock waves. It is demonstrated that the strain rates for the brain 1 are in the range of 36 to 241 s− , approximately 1.9 and 0.86 times the resulting head acceleration under impacts and blast scenarios, respectively. The skull was found to experience a rate in the range 1 of 14 to 182 s− , approximately 0.7 and 0.43 times the head acceleration corresponding to impact and blast cases. -

Corpora Amylacea Act As Containers That Remove Waste Products from the Brain
Corpora amylacea act as containers that remove waste products from the brain Marta Ribaa,b,c,1, Elisabet Augéa,b,c,1, Joan Campo-Sabariza,d, David Moral-Antera,d, Laura Molina-Porcele, Teresa Ximelise, Ruth Ferrera,d, Raquel Martín-Venegasa,d, Carme Pelegría,b,c,2, and Jordi Vilaplanaa,b,c,2 aSecció de Fisiologia, Departament de Bioquímica i Fisiologia, Universitat de Barcelona, 08028 Barcelona, Spain; bInstitut de Neurociències, Universitat de Barcelona, 08035 Barcelona, Spain; cCentros de Biomedicina en Red de Enfermedades Neurodegenerativas (CIBERNED), 28031 Madrid, Spain; dInstitut de Recerca en Nutrició i Seguretat Alimentàries (INSA-UB), Universitat de Barcelona, 08291 Barcelona, Spain; and eNeurological Tissue Bank of the Biobanc- Hospital Clinic-Institut d’Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS), 08036 Barcelona, Spain Edited by Lawrence Steinman, Stanford University School of Medicine, Stanford, CA, and approved November 5, 2019 (received for review August 8, 2019) Corpora amylacea (CA) in the human brain are granular bodies 6). Nonetheless, we observed that CA contain glycogen synthase formed by polyglucosan aggregates that amass waste products of (GS), an indispensable enzyme for polyglucosan formation, and different origins. They are generated by astrocytes, mainly during also ubiquitin and protein p62, both associated with processes of aging and neurodegenerative conditions, and are located predom- elimination of waste substances (5). The relationship between CA inantly in periventricular and subpial regions. This study shows that and waste substances is recurrent in the literature. Already in 1999, CA are released from these regions to the cerebrospinal fluid and after a detailed and complete review, Cavanagh indicated that “CA are present in the cervical lymph nodes, into which cerebrospinal functions seem to be directed towards trapping and sequestration fluid drains through the meningeal lymphatic system. -

A Cellular Atlas of the Developing Meninges Reveals Meningeal Fibroblast Diversity and Function
bioRxiv preprint doi: https://doi.org/10.1101/648642; this version posted May 24, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 2 3 4 Title: A cellular atlas of the developing meninges reveals meningeal fibroblast diversity and function 5 6 Authors: John DeSisto1,2,3,, Rebecca O’Rourke2, Stephanie Bonney1,3, Hannah E. Jones1,3, Fabien 7 Guimiot4, Kenneth L. Jones2 and Julie A. Siegenthaler1,3,5 8 9 1Department of Pediatrics Section of Developmental Biology, 2Department of Pediatrics Section of 10 Section of Hematology, Oncology, Bone Marrow Transplant, 3Cell Biology, Stem Cells and Development 11 Graduate Program, University of Colorado, Anschutz Medical Campus, Aurora, CO 80045 USA, 12 4INSERM UMR 1141, Hôpital Robert Debré, 75019 Paris, France. 13 14 5Corresponding Author: 15 Julie A. Siegenthaler, PhD 16 University of Colorado, School of Medicine 17 Department of Pediatrics 18 12800 East 19th Ave MS-8313 19 Aurora, CO 80045 USA 20 Telephone #: 303-724-3123 21 E-mail: [email protected] 22 23 Key words (3-6 words): brain development, meninges, pial basement membrane, retinoic acid, human 24 meninges bioRxiv preprint doi: https://doi.org/10.1101/648642; this version posted May 24, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 25 Abstract 26 The meninges, a multilayered structure that encases the CNS, is composed mostly of fibroblasts, 27 along with vascular and immune cells. -

Adaptive Immune Systems
Immunology 101 (for the Non-Immunologist) Abhinav Deol, MD Assistant Professor of Oncology Wayne State University/ Karmanos Cancer Institute, Detroit MI Presentation originally prepared and presented by Stephen Shiao MD, PhD Department of Radiation Oncology Cedars-Sinai Medical Center Disclosures Bristol-Myers Squibb – Contracted Research What is the immune system? A network of proteins, cells, tissues and organs all coordinated for one purpose: to defend one organism from another It is an infinitely adaptable system to combat the complex and endless variety of pathogens it must address Outline Structure of the immune system Anatomy of an immune response Role of the immune system in disease: infection, cancer and autoimmunity Organs of the Immune System Major organs of the immune system 1. Bone marrow – production of immune cells 2. Thymus – education of immune cells 3. Lymph Nodes – where an immune response is produced 4. Spleen – dual role for immune responses (especially antibody production) and cell recycling Origins of the Immune System B-Cell B-Cell Self-Renewing Common Progenitor Natural Killer Lymphoid Cell Progenitor Thymic T-Cell Selection Hematopoetic T-Cell Stem Cell Progenitor Dendritic Cell Myeloid Progenitor Granulocyte/M Macrophage onocyte Progenitor The Immune Response: The Art of War “Know your enemy and know yourself and you can fight a hundred battles without disaster.” -Sun Tzu, The Art of War Immunity: Two Systems and Their Key Players Adaptive Immunity Innate Immunity Dendritic cells (DC) B cells Phagocytes (Macrophages, Neutrophils) Natural Killer (NK) Cells T cells Dendritic Cells: “Commanders-in-Chief” • Function: Serve as the gateway between the innate and adaptive immune systems. -

Human Anatomy As Related to Tumor Formation Book Four
SEER Program Self Instructional Manual for Cancer Registrars Human Anatomy as Related to Tumor Formation Book Four Second Edition U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service National Institutesof Health SEER PROGRAM SELF-INSTRUCTIONAL MANUAL FOR CANCER REGISTRARS Book 4 - Human Anatomy as Related to Tumor Formation Second Edition Prepared by: SEER Program Cancer Statistics Branch National Cancer Institute Editor in Chief: Evelyn M. Shambaugh, M.A., CTR Cancer Statistics Branch National Cancer Institute Assisted by Self-Instructional Manual Committee: Dr. Robert F. Ryan, Emeritus Professor of Surgery Tulane University School of Medicine New Orleans, Louisiana Mildred A. Weiss Los Angeles, California Mary A. Kruse Bethesda, Maryland Jean Cicero, ART, CTR Health Data Systems Professional Services Riverdale, Maryland Pat Kenny Medical Illustrator for Division of Research Services National Institutes of Health CONTENTS BOOK 4: HUMAN ANATOMY AS RELATED TO TUMOR FORMATION Page Section A--Objectives and Content of Book 4 ............................... 1 Section B--Terms Used to Indicate Body Location and Position .................. 5 Section C--The Integumentary System ..................................... 19 Section D--The Lymphatic System ....................................... 51 Section E--The Cardiovascular System ..................................... 97 Section F--The Respiratory System ....................................... 129 Section G--The Digestive System ......................................... 163 Section -
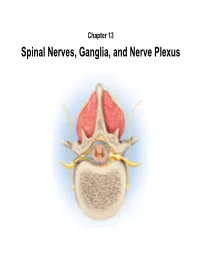
Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves
Chapter 13 Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves Posterior Spinous process of vertebra Posterior root Deep muscles of back Posterior ramus Spinal cord Transverse process of vertebra Posterior root ganglion Spinal nerve Anterior ramus Meningeal branch Communicating rami Anterior root Vertebral body Sympathetic ganglion Anterior General Anatomy of Nerves and Ganglia • Spinal cord communicates with the rest of the body by way of spinal nerves • nerve = a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue – mixed nerves contain both afferent (sensory) and efferent (motor) fibers – composed of thousands of fibers carrying currents in opposite directions Anatomy of a Nerve Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Epineurium Perineurium Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Endoneurium Nerve Rootlets fiber Posterior root Fascicle Posterior root ganglion Anterior Blood root vessels Spinal nerve (b) Copyright by R.G. Kessel and R.H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, 1979, W.H. Freeman, All rights reserved Blood vessels Fascicle Epineurium Perineurium Unmyelinated nerve fibers Myelinated nerve fibers (a) Endoneurium Myelin General Anatomy of Nerves and Ganglia • nerves of peripheral nervous system are ensheathed in Schwann cells – forms neurilemma and often a myelin sheath around the axon – external to neurilemma, each fiber is surrounded by -

The Peripheral Nervous System
The Peripheral Nervous System Dr. Ali Ebneshahidi Peripheral Nervous System (PNS) – Consists of 12 pairs of cranial nerves and 31 pairs of spinal nerves. – Serves as a critical link between the body and the central nervous system. – peripheral nerves contain an outermost layer of fibrous connective tissue called epineurium which surrounds a thinner layer of fibrous connective tissue called perineurium (surrounds the bundles of nerve or fascicles). Individual nerve fibers within the nerve are surrounded by loose connective tissue called endoneurium. Cranial Nerves Cranial nerves are direct extensions of the brain. Only Nerve I (olfactory) originates from the cerebrum, the remaining 11 pairs originate from the brain stem. Nerve I (Olfactory)- for the sense of smell (sensory). Nerve II (Optic)- for the sense of vision (sensory). Nerve III (Oculomotor)- for controlling muscles and accessory structures of the eyes ( primarily motor). Nerve IV (Trochlear)- for controlling muscles of the eyes (primarily motor). Nerve V (Trigeminal)- for controlling muscles of the eyes, upper and lower jaws and tear glands (mixed). Nerve VI (Abducens)- for controlling muscles that move the eye (primarily motor). Nerve VII (Facial) – for the sense of taste and controlling facial muscles, tear glands and salivary glands (mixed). Nerve VIII (Vestibulocochlear)- for the senses of hearing and equilibrium (sensory). Nerve IX (Glossopharyngeal)- for controlling muscles in the pharynx and to control salivary glands (mixed). Nerve X (Vagus)- for controlling muscles used in speech, swallowing, and the digestive tract, and controls cardiac and smooth muscles (mixed). Nerve XI (Accessory)- for controlling muscles of soft palate, pharynx and larynx (primarily motor). Nerve XII (Hypoglossal) for controlling muscles that move the tongue ( primarily motor). -
PN1 (Midha) Microanatomy of Peripheral Nerves-Part1.Pdf
Peripheral Nerve Basics • Consist of processes of cell bodies found in the DRG, anterior horn, and autonomic ganglia • Organized by several distinct connective tissue layers – the epineurium, perineurium, and endoneurium • Vascular supply provided by the vasa nervorum Peripheral Nerve Basics • Neuronal processes bound into fascicles by perineurium • Fascicles bound into nerves by epineurium • Endoneurium is a division of the perineurium which form thin layers of connective tissue surrounding neuronal fibers in a fascicle Sural nerve in cross-section Epineurium • Loose areolar tissue with sparse, longitudinally-oriented collagen fibers • Some elastic fibers where epineurium abuts perineurium • Able to accommodate a significant amount of nerve stretching and movement • Increases in thickness where nerves cross joints • Constitutes an increasing proportion of nerves as they increase in size • Epineurial fat helps cushion nerves from compressive injury • Decreased epineurial fat found in patients with diabetes Perineurium • Cellular component composed of laminated fibroblasts of up to 15 layers in thickness which are bounded by a basal lamina • Semi-permeable: inner lamellae have tight junctions, providing a barrier to intercellular transport of macromolecules – Tight junctions can be loosened with topical anaesthetics and with osmotic change Perineurium • Exhibits a slightly positive internal pressure – Fascicular contents herniate upon perineurial injury • Under tension longitudinally – Nerve segment shortens upon transection – may complicate surgical repair as nerve can be stretched only approximately 10% before being inhibited by collagen Endoneurium • Intrafascicular connective tissue consisting of a collagenous matrix in the interstitial space • Develops into partitions of dense connective tissue between diverging fascicles and eventually becomes perineurium when the fascicles separate • Collagen fibers are longitudinally-oriented and run along nerve fibers and capillaries.