Trivalent Phosphorus and Phosphines As Components of Biochemistry in Anoxic Environments
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Use of Phosphate Solubilizing Bacteria to Leach Rare Earth Elements from Monazite-Bearing Ore
Minerals 2015, 5, 189-202; doi:10.3390/min5020189 OPEN ACCESS minerals ISSN 2075-163X www.mdpi.com/journal/minerals Article Use of Phosphate Solubilizing Bacteria to Leach Rare Earth Elements from Monazite-Bearing Ore Doyun Shin 1,2,*, Jiwoong Kim 1, Byung-su Kim 1,2, Jinki Jeong 1,2 and Jae-chun Lee 1,2 1 Mineral Resources Resource Division, Korea Institute of Geoscience and Mineral Resources (KIGAM), Gwahangno 124, Yuseong-gu, Daejeon 305-350, Korea; E-Mails: [email protected] (J.K.); [email protected] (B.K.); [email protected] (J.J.); [email protected] (J.L.) 2 Department of Resource Recycling Engineering, Korea University of Science and Technology, Gajeongno 217, Yuseong-gu, Daejeon 305-350, Korea * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +82-42-868-3616. Academic Editor: Anna H. Kaksonen Received: 8 January 2015 / Accepted: 27 March 2015 / Published: 2 April 2015 Abstract: In the present study, the feasibility to use phosphate solubilizing bacteria (PSB) to develop a biological leaching process of rare earth elements (REE) from monazite-bearing ore was determined. To predict the REE leaching capacity of bacteria, the phosphate solubilizing abilities of 10 species of PSB were determined by halo zone formation on Reyes minimal agar media supplemented with bromo cresol green together with a phosphate solubilization test in Reyes minimal liquid media as the screening studies. Calcium phosphate was used as a model mineral phosphate. Among the test PSB strains, Pseudomonas fluorescens, P. putida, P. rhizosphaerae, Mesorhizobium ciceri, Bacillus megaterium, and Acetobacter aceti formed halo zones, with the zone of A. -

Prebiological Evolution and the Metabolic Origins of Life
Prebiological Evolution and the Andrew J. Pratt* Metabolic Origins of Life University of Canterbury Keywords Abiogenesis, origin of life, metabolism, hydrothermal, iron Abstract The chemoton model of cells posits three subsystems: metabolism, compartmentalization, and information. A specific model for the prebiological evolution of a reproducing system with rudimentary versions of these three interdependent subsystems is presented. This is based on the initial emergence and reproduction of autocatalytic networks in hydrothermal microcompartments containing iron sulfide. The driving force for life was catalysis of the dissipation of the intrinsic redox gradient of the planet. The codependence of life on iron and phosphate provides chemical constraints on the ordering of prebiological evolution. The initial protometabolism was based on positive feedback loops associated with in situ carbon fixation in which the initial protometabolites modified the catalytic capacity and mobility of metal-based catalysts, especially iron-sulfur centers. A number of selection mechanisms, including catalytic efficiency and specificity, hydrolytic stability, and selective solubilization, are proposed as key determinants for autocatalytic reproduction exploited in protometabolic evolution. This evolutionary process led from autocatalytic networks within preexisting compartments to discrete, reproducing, mobile vesicular protocells with the capacity to use soluble sugar phosphates and hence the opportunity to develop nucleic acids. Fidelity of information transfer in the reproduction of these increasingly complex autocatalytic networks is a key selection pressure in prebiological evolution that eventually leads to the selection of nucleic acids as a digital information subsystem and hence the emergence of fully functional chemotons capable of Darwinian evolution. 1 Introduction: Chemoton Subsystems and Evolutionary Pathways Living cells are autocatalytic entities that harness redox energy via the selective catalysis of biochemical transformations. -

Discovery, Metabolism and Functions of NAD and NADP
Coenzymes Features Discovery, metabolism and functions of NAD Downloaded from http://portlandpress.com/biochemist/article-pdf/37/1/9/3189/bio037010009.pdf by guest on 01 October 2021 and NADP Magali R. VanLinden, Renate Hvidsten Skoge and Mathias Ziegler (University of Bergen, Norway) Nicotinamide adenine dinucleotide (NAD) and nicotinamide adenine dinucleotide phosphate (NADP) are two major players in metabolism as they participate as electron carriers in a multitude of redox reactions. Moreover, they act in life and death decisions on a cellular level in all known life forms. NAD and NADP both exist in two states; the oxidized forms are characterized by a positive charge on the nicotinamide (Nam) moiety, denoted NAD+ and NADP+ respectively. The reduced forms are denoted NADH and NADPH (Figure 1). The independent discoveries of NAD(P) as vitamins and co-enzymes Vitamin B3 is a collective term for the two NAD(P) precursors nicotinic acid (NA) (also referred to as niacin) and Nam, as well as their corresponding ribosides. Niacin and Nam were found to be essential nutrients after severe outbreaks of pellagra (originally thought to be a new pestilence) in 18th Century Europe. Large outbreaks of pellagra occurred in North America in the early 20th Century. Pellagra (from Italian: pelle = skin; agra = sour) is characterized by dermatitis, diarrhoea, dementia and ultimately death, and was widespread in the hundreds of thousands of poor people living on a diet mostly composed of corn or maize flour. Although successfully consumed by American Indians for centuries, degerminated maize does not contain niacin in a bioavailable form. The crucial difference was that Figure 1. -

Sulfonyl-Containing Nucleoside Phosphotriesters And
Sulfonyl-Containing Nucleoside Phosphotriesters and Phosphoramidates as Novel Anticancer Prodrugs of 5-Fluoro-2´-Deoxyuridine-5´- Monophosphate (FdUMP) Yuan-Wan Sun, Kun-Ming Chen, and Chul-Hoon Kwon†,* †Department of Pharmaceutical Sciences, College of Pharmacy and Allied Health Professions, St. John’s University, Jamaica, New York 11439 * To whom correspondence should be addressed. Department of Pharmaceutical Sciences, College of Pharmacy and Allied Health Professions, St. John’s University, 8000 Utopia parkway, Jamaica, NY 11439. Tel: (718)-990-5214, fax: (718)-990-6551, e-mail: [email protected]. Abstract A series of sulfonyl-containing 5-fluoro-2´-deoxyuridine (FdU) phosphotriester and phosphoramidate analogues were designed and synthesized as anticancer prodrugs of FdUMP. Stability studies have demonstrated that these compounds underwent pH dependent β-elimination to liberate the corresponding nucleotide species with half-lives in the range of 0.33 to 12.23 h under model physiological conditions in 0.1M phosphate buffer at pH 7.4 and 37 °C. Acceleration of the elimination was observed in the presence of human plasma. Compounds with FdUMP moiety (4-9) were considerably more potent than those without (1-3) as well as 5-fluorouracil (5-FU) against Chinese hamster lung fibroblasts (V-79 cells) in vitro. Addition of thymidine (10 µM) reversed the growth inhibition activities of only 5-FU and the compounds with FdUMP moiety, but had no effect on those without. These results suggested a mechanism of action of the prodrugs involving the intracellular release of FdUMP. Introduction 5-Fluoro-2´-deoxyuridine-5´-monophosphate (FdUMP) is the major metabolite responsible for the anticancer activity of 5-FU (Chart 1). -

Chemical Warfare Agent (CWA) Identification Overview
Physicians for Human Rights Chemical Warfare Agent (CWA) Identification Overview Chemical Warfare Agent Identification Fact Sheet Series Table of Contents This Chemical Warfare Agent (CWA) Identification Fact Sheet is part 2 Physical Properties of a Physicians for Human Rights (PHR) series designed to fill a gap in 2 VX (Nerve Agent) 2 Sarin (Nerve Agent) knowledge among medical first responders to possible CWA attacks. 2 Tabun (Nerve Agent) This document in particular outlines differences between a select 2 BZ (Incapacitating Agent) group of vesicants and nerve agents, the deployment of which would 2 Mustard Gas (Vesicant) necessitate emergency medical treatment and documentation. 3 Collecting Samples to Test for Exposure 4 Protection PHR hopes that, by referencing these fact sheets, medical professionals 5 Symptoms may be able to correctly diagnose, treat, and document evidence of 6 Differential Diagnosis exposure to CWAs. Information in this fact sheet has been compiled from 8 Decontimanation 9 Treatment publicly available sources. 9 Abbreviations A series of detailed CWA fact sheets outlining in detail those properties and treatment regimes unique to each CWA is available at physiciansforhumanrights.org/training/chemical-weapons. phr.org Chemical Warfare Agent (CWA) Identification Overview 1 Collect urine samples, and blood and hair samples if possible, immediately after exposure Physical Properties VX • A lethal dose (10 mg) of VX, absorbed through the skin, can kill within minutes (Nerve Agent) • Can remain in environment for weeks -
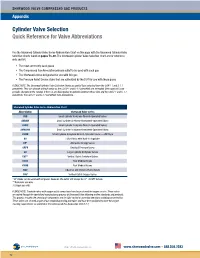
Cylinder Valve Selection Quick Reference for Valve Abbreviations
SHERWOOD VALVE COMPRESSED GAS PRODUCTS Appendix Cylinder Valve Selection Quick Reference for Valve Abbreviations Use the Sherwood Cylinder Valve Series Abbreviation Chart on this page with the Sherwood Cylinder Valve Selection Charts found on pages 73–80. The Sherwood Cylinder Valve Selection Chart are for reference only and list: • The most commonly used gases • The Compressed Gas Association primary outlet to be used with each gas • The Sherwood valves designated for use with this gas • The Pressure Relief Device styles that are authorized by the DOT for use with these gases PLEASE NOTE: The Sherwood Cylinder Valve Selection Charts are partial lists extracted from the CGA V-1 and S-1.1 pamphlets. They can change without notice as the CGA V-1 and S-1.1 pamphlets are amended. Sherwood will issue periodic changes to the catalog. If there is any discrepancy or question between these lists and the CGA V-1 and S-1.1 pamphlets, the CGA V-1 and S-1.1 pamphlets take precedence. Sherwood Cylinder Valve Series Abbreviation Chart Abbreviation Sherwood Valve Series AVB Small Cylinder Acetylene Wrench-Operated Valves AVBHW Small Cylinder Acetylene Handwheel-Operated Valves AVMC Small Cylinder Acetylene Wrench-Operated Valves AVMCHW Small Cylinder Acetylene Handwheel-Operated Valves AVWB Small Cylinder Acetylene Wrench-Operated Valves — WB Style BV Hi/Lo Valves with Built-in Regulator DF* Alternative Energy Valves GRPV Residual Pressure Valves GV Large Cylinder Acetylene Valves GVT** Vertical Outlet Acetylene Valves KVAB Post Medical Valves KVMB Post Medical Valves NGV Industrial and Chrome-Plated Valves YVB† Vertical Outlet Oxygen Valves 1 * DF Valves can be used with all gases; however, the outlet will always be ⁄4"–18 NPT female. -

A Comparative Study of the Formation of Aromatics In
A COMPARATIVE STUDY OF THE FORMATION OF AROMATICS IN RICH METHANE FLAMES DOPED BY UNSATURATED COMPOUNDS H.A. GUENICHE, J. BIET, P.A. GLAUDE, R. FOURNET, F. BATTIN-LECLERC* Département de Chimie-Physique des Réactions, Nancy Université, CNRS, 1 rue Grandville, BP 20451, 54001 NANCY Cedex, France * E-mail : [email protected] ; Tel.: 33 3 83 17 51 25 , Fax : 33 3 83 37 81 20 ABSTRACT For a better modeling of the importance of the different channels leading to the first aromatic ring, we have compared the structures of laminar rich premixed methane flames doped with several unsaturated hydrocarbons: allene and propyne, because they are precursors of propargyl radicals which are well known as having an important role in forming benzene, 1,3-butadiene to put in evidence a possible production of benzene due to reactions of C4 compounds, and, finally, cyclopentene which is a source of cyclopentadienylmethylene radicals which in turn are expected to easily isomerizes to give benzene. These flames have been stabilized on a burner at a pressure of 6.7 kPa (50 Torr) using argon as dilutant, for equivalence ratios (φ) from 1.55 to 1.79. A unique mechanism, including the formation and decomposition of benzene and toluene, has been used to model the oxidation of allene, propyne, 1,3-butadiene and cyclopentene. The main reaction pathways of aromatics formation have been derived from reaction rate and sensitivity analyses and have been compared for the three types of additives. These combined analyses and comparisons can only been performed when a unique mechanism is available for all the studied additives. -

MMM #278 Since December 1986 SEPTEMBER 2014 – P 1
MMM #278 Since December 1986 SEPTEMBER 2014 – p 1 Two worlds of the Inner Solar System that present enormous challenges for human visitors, explorers, and settlers Feature Articles: 2 In Focus: Venus & Mercury: Why Limit Human Frontiers to Moon and Mars? 3 Venus: The Sources of Radical Transformation are already “on Location” 5 Mercury: Discovery of a hidden Settlement Sweet Spot; “Location, Location, Location” 7 Moon Base Costs: Dave Dietzler Below: Previous Articles about Venus have focused on “Aerostat stations” high above the surface, where air pressure and temperatures are human-friendly Above left: a Venus aerostat station - right: at an altitude where temperatures and pressures are benign For past articles, Visit http://www.moonsociety.org/publications/mmm_classics/ or /mmm_themes/ MMM #278 Since December 1986 SEPTEMBER 2014 – p 2 About Moon Miners’ Manifesto - “The Moon - it’s not Earth, but it’s Earth’s!” • MMM’s VISION: “expanding the human economy through of-planet resources”; early heavy reliance on Lunar materials; early use of Mars system and asteroid resources; and permanent settlements supporting this economy. • MMM’s MISSION: to encourage “spin-up” entrepreneurial development of the novel technologies needed and promote the economic-environmental rationale of space and lunar settlement. • Moon Miners’ Manifesto CLASSICS: The non-time-sensitive articles and editorials of MMM’s first twenty years plus have been re-edited, reillustrated, and republished in 23 PDF format volumes, for free downloading from this location: http://www.MoonSociety.org/publications/mmm_classics/ • MMM THEME Issues: 14 collections of articles according to themes: ..../publications/mmm_themes/ • MMM Glossary: new terms, old terms/new meanings: www.moonsociety.org/publications/m3glossary.html • MMM retains its editorial independence and serves many groups, each with its own philosophy, agenda, and programs. -

High Purity/Recovery Separation of Propylene from Propyne Using Anion Pillared Metal-Organic Framework: Application of Vacuum Swing Adsorption (VSA)
energies Article High Purity/Recovery Separation of Propylene from Propyne Using Anion Pillared Metal-Organic Framework: Application of Vacuum Swing Adsorption (VSA) Majeda Khraisheh 1, Fares AlMomani 1,* and Gavin Walker 2 1 Department of Chemical Engineering, College of Engineering, Qatar University, Doha P.O. Box 2713, Qatar; [email protected] 2 Department of Chemical Sciences, SSPC, Bernal Institute, University of Limerick, V94 T9PX Limerick, Ireland; [email protected] * Correspondence: [email protected]; Tel.: +974-4403-4140 Abstract: Propylene is one of the world’s most important basic olefin raw material used in the produc- tion of a vast array of polymers and other chemicals. The need for high purity grade of propylene is essential and traditionally achieved by the very energy-intensive cryogenic separation. In this study, 2− a pillared inorganic anion SIF6 was used as a highly selective C3H4 due to the square grid pyrazine- based structure. Single gas adsorption revealed a very high C3H4 uptake value (3.32, 3.12, 2.97 and 2.43 mmol·g−1 at 300, 320, 340 and 360 K, respectively). The values for propylene for the same tempera- tures were 2.73, 2.64, 2.31 and 1.84 mmol·g−1, respectively. Experimental results were obtained for the two gases fitted using Langmuir and Toth models. The former had a varied degree of representation of the system with a better presentation of the adsorption of the propylene compared to the propyne system. The Toth model regression offered a better fit of the experimental data over the entire range of pressures. -

Section 2. Hazards Identification OSHA/HCS Status : This Material Is Considered Hazardous by the OSHA Hazard Communication Standard (29 CFR 1910.1200)
SAFETY DATA SHEET Nonflammable Gas Mixture: Arsine / Helium / Phosgene Section 1. Identification GHS product identifier : Nonflammable Gas Mixture: Arsine / Helium / Phosgene Other means of : Not available. identification Product use : Synthetic/Analytical chemistry. SDS # : 006605 Supplier's details : Airgas USA, LLC and its affiliates 259 North Radnor-Chester Road Suite 100 Radnor, PA 19087-5283 1-610-687-5253 24-hour telephone : 1-866-734-3438 Section 2. Hazards identification OSHA/HCS status : This material is considered hazardous by the OSHA Hazard Communication Standard (29 CFR 1910.1200). Classification of the : GASES UNDER PRESSURE - Compressed gas substance or mixture ACUTE TOXICITY (inhalation) - Category 4 GHS label elements Hazard pictograms : Signal word : Warning Hazard statements : Contains gas under pressure; may explode if heated. May displace oxygen and cause rapid suffocation. Harmful if inhaled. Precautionary statements General : Read and follow all Safety Data Sheets (SDS’S) before use. Read label before use. Keep out of reach of children. If medical advice is needed, have product container or label at hand. Close valve after each use and when empty. Use equipment rated for cylinder pressure. Do not open valve until connected to equipment prepared for use. Use a back flow preventative device in the piping. Use only equipment of compatible materials of construction. Prevention : Use only outdoors or in a well-ventilated area. Avoid breathing gas. Response : IF INHALED: Remove person to fresh air and keep comfortable for breathing. Call a POISON CENTER or physician if you feel unwell. Storage : Protect from sunlight when ambient temperature exceeds 52°C/125°F. Store in a well- ventilated place. -

Modern Chemical Weapons
Modern Chemical Weapons Modern Chemical Weapons causes serious diseases like cancer and serious birth defects in newly born Large scale chemical weapons were children. first used in World War One and have been used ever since. About 100 years ago Modern warfare has developed significantly since the early 20th century Early chemical weapons being used but the use of toxic chemicals to kill around a hundred years ago included: and badly injure is still very much in use tear gas, chlorine gas, mustard gas today. There have been reports of and phosgene gas. Since then, some chemical weapon attacks in Syria of the same chemicals have been during 2016. Chemical weapons have used in more modern warfare, but also been the choice of terrorists other new chemical weapons have because they are not very expensive also been developed. and need very little specialist knowledge to use them. These Chlorine gas (Cl2) weapons can cause a lot of causalities as well as fatalities, but also There have been reports of many spread panic and fear. chlorine gas attacks in Syria since 2013. It is a yellow-green gas which has a very distinctive smell like bleach. However, it does not last very long and therefore it is sometimes very difficult to prove it has been used during an attack. Victims would feel irritation of the eyes, nose, throat and lungs when they inhale it in large enough quantities. In even larger quantities it can cause the death of a person by suffocation. Mustard gas (C4H8Cl2S) There are reports by the United Nations (UN) of terrorist groups using Mustard Agent Orange (mixture of gas during chemical attacks in Syria. -

First Life in Primordial-Planet Oceans: the Biological Big Bang Gibson/Wickramasinghe/Schild First Life in Primordial-Planet Oceans: the Biological Big Bang
Journal of Cosmology First life in primordial-planet oceans: the biological big bang Gibson/Wickramasinghe/Schild First life in primordial-planet oceans: the biological big bang Carl H. Gibson 1,2 1 University of California San Diego, La Jolla, CA 92093-0411, USA [email protected], http://sdcc3.ucsd.edu/~ir118 N. Chandra Wickramasinghe3,4 3Cardiff Centre for Astrobiology, 24 Llwynypia Road, Lisvane, Cardiff CF14 0SY [email protected] Rudolph E. Schild5,6 5 Center for Astrophysics, 60 Garden Street, Cambridge, MA 02138, USA 6 [email protected] Abstract: A scenario is presented for the formation of first life in the universe based on hydro- gravitational-dynamics (HGD) cosmology. From HGD, the dark matter of galaxies is H-He gas dominated planets (primordial-fog-particle PFPs) in million solar mass clumps (protoglobularstar- cluster PGCs), which formed at the plasma to gas transition temperature 3000 K. Stars result from mergers of these hot-gas-planets. Over-accretion causes stars to explode as supernovae that scatter life-chemicals (C, N, O, P, S, Ca, Fe etc.) to other planets in PGC clumps and beyond. These chemicals were first collected gravitationally by merging PFPs to form H-saturated, high-pressure, dense oceans of critical-temperature 647 K water over iron-nickel cores at ~ 2 Myr. Stardust fertil- izes the formation of first life in a cosmic hot-ocean soup kitchen comprised of all planets and their moons in meteoric communication, > 10100 kg in total. Ocean freezing at 273 K slows this biologi- cal big bang at ~ 8 Myr. HGD cosmology confirms that the evolving seeds of life are scattered on intergalactic scales by Hoyle-Wickramasinghe cometary panspermia.