Quality of Main Types of Hunted Red Deer Meat Obtained in Spain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
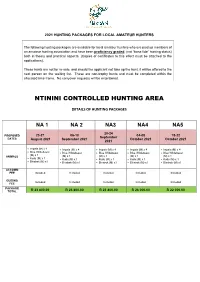
Ntinini Controlled Hunting Area
2021 HUNTING PACKAGES FOR LOCAL AMATEUR HUNTERS The following hunting packages are available for local amateur hunters who are paid up members of an amateur hunting association and have been proficiency graded, (not “bona fide” hunting status) both in theory and practical aspects. (Copies of certificates to this effect must be attached to the applications). These hunts are not for re-sale, and should the applicant not take up the hunt, it will be offered to the next person on the waiting list. These are non-trophy hunts and must be completed within the allocated time frame. No carryover requests will be entertained. NTININI CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES NA 1 NA 2 NA3 NA4 NA5 20-24 23-27 06-10 04-08 18-22 PROPOSED September DATES August 2021 September 2021 October 2021 October 2021 2021 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest (M) x 1 ANIMALS (M) x 1 (M) x 1 (M) x 1 (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 ACCOMM FEE Included Included Included Included Included GUIDING Included Included Included Included Included FEE PACKAGE TOTAL R 23 800.00 R 23 800.00 R 23 800.00 R 26 000.00 R 22 000.00 ITHALA CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES IGRA 1 IGRA 2 IGRA 3 IGRA 4 PROPOSED 11-16 02-07 16-21 06-11 DATES June 2021 July 2021 July 2021 August 2021 • Impala -

Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout Has in THIS ISSUE Dbeen Awarded the President’S Cup Letter from the President
DSC NEWSLETTER VOLUME 32,Camp ISSUE 5 TalkJUNE 2019 Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout has IN THIS ISSUE Dbeen awarded the President’s Cup Letter from the President .....................1 for her pending world record East African DSC Foundation .....................................2 Defassa Waterbuck. Awards Night Results ...........................4 DSC’s April Monthly Meeting brings Industry News ........................................8 members together to celebrate the annual Chapter News .........................................9 Trophy and Photo Award presentation. Capstick Award ....................................10 This year, there were over 150 entries for Dove Hunt ..............................................12 the Trophy Awards, spanning 22 countries Obituary ..................................................14 and almost 100 different species. Membership Drive ...............................14 As photos of all the entries played Kid Fish ....................................................16 during cocktail hour, the room was Wine Pairing Dinner ............................16 abuzz with stories of all the incredible Traveler’s Advisory ..............................17 adventures experienced – ibex in Spain, Hotel Block for Heritage ....................19 scenic helicopter rides over the Northwest Big Bore Shoot .....................................20 Territories, puku in Zambia. CIC International Conference ..........22 In determining the winners, the judges DSC Publications Update -

An Analysis of the Phylogenetic Relationship of Thai Cervids Inferred from Nucleotide Sequences of Protein Kinase C Iota (PRKCI) Intron
Kasetsart J. (Nat. Sci.) 43 : 709 - 719 (2009) An Analysis of the Phylogenetic Relationship of Thai Cervids Inferred from Nucleotide Sequences of Protein Kinase C Iota (PRKCI) Intron Kanita Ouithavon1, Naris Bhumpakphan2, Jessada Denduangboripant3, Boripat Siriaroonrat4 and Savitr Trakulnaleamsai5* ABSTRACT The phylogenetic relationship of five Thai cervid species (n=21) and four spotted deer (Axis axis, n=4), was determined based on nucleotide sequences of the intron region of the protein kinase C iota (PRKCI) gene. Blood samples were collected from seven captive breeding centers in Thailand from which whole genomic DNA was extracted. Intron1 sequences of the PRKCI nuclear gene were amplified by a polymerase chain reaction, using L748 and U26 primers. Approximately 552 base pairs of all amplified fragments were analyzed using the neighbor-joining distance matrix method, and 19 parsimony- informative sites were analyzed using the maximum parsimony approach. Phylogenetic analyses using the subfamily Muntiacinae as outgroups for tree rooting indicated similar topologies for both phylogenetic trees, clearly showing a separation of three distinct genera of Thai cervids: Muntiacus, Cervus, and Axis. The study also found that a phylogenetic relationship within the genus Axis would be monophyletic if both spotted deer and hog deer were included. Hog deer have been conventionally classified in the genus Cervus (Cervus porcinus), but this finding supports a recommendation to reclassify hog deer in the genus Axis. Key words: Thailand, the family Cervidae, protein kinase C iota (PRKCI) intron, phylogenetic tree, taxonomy INTRODUCTION 1987; Gentry, 1994). Cervids, or what is commonly called “deer,” are mostly characterized The family Cervidae is one of the most by antlers with a bony inner core and velvet skin specious families of artiodactyls, with an extensive cover. -

Influence of Common Eland (Taurotragus Oryx) Meat Composition on Its Further Technological Processing
CZECH UNIVERSITY OF LIFE SCIENCES PRAGUE Faculty of Tropical AgriSciences Department of Animal Science and Food Processing Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing DISSERTATION THESIS Prague 2018 Author: Supervisor: Ing. et Ing. Petr Kolbábek prof. MVDr. Daniela Lukešová, CSc. Co-supervisors: Ing. Radim Kotrba, Ph.D. Ing. Ludmila Prokůpková, Ph.D. Declaration I hereby declare that I have done this thesis entitled “Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing” independently, all texts in this thesis are original, and all the sources have been quoted and acknowledged by means of complete references and according to Citation rules of the FTA. In Prague 5th October 2018 ………..………………… Acknowledgements I would like to express my deep gratitude to prof. MVDr. Daniela Lukešová CSc., Ing. Radim Kotrba, Ph.D. and Ing. Ludmila Prokůpková, Ph.D., and doc. Ing. Lenka Kouřimská, Ph.D., my research supervisors, for their patient guidance, enthusiastic encouragement and useful critiques of this research work. I am very gratefull to Ing. Petra Maxová and Ing. Eva Kůtová for their valuable help during the research. I am also gratefull to Mr. Petr Beluš, who works as a keeper of elands in Lány, Mrs. Blanka Dvořáková, technician in the laboratory of meat science. My deep acknowledgement belongs to Ing. Radek Stibor and Mr. Josef Hora, skilled butchers from the slaughterhouse in Prague – Uhříněves and to JUDr. Pavel Jirkovský, expert marksman, who shot the animals. I am very gratefull to the experts from the Natura Food Additives, joint-stock company and from the Alimpex-maso, Inc. -

A National Park in Northern Rhodesia 15
A National Park in Northern Rhodesia 15 A NATIONAL PARK IN NORTHERN RHODESIA By a Proclamation dated 20th April, 1950, the Governor of Northern Rhodesia has established a National Park, to be known as the Kafue National Park. This new park covers some 8,650 square miles roughly in the central Kafue basin, between latitudes 14° and 16° 40' S. It contains a wide range of country from the comparatively drv sandy lands of the south to the big rivers, swamps, and heavy timber of the northern section. The magnificent Kafue River dominates the whole central portion, adding scenic beauty to the attraction of wild life. The park contains representatives of most species of the fauna of Northern Rhodesia. Primates are represented by the Rhodesian Baboon, the Vervet Monkey, and the Greater and Lesser Night-Apes. There are Elephant and Black Rhinoceros, Buffalo, Lion, Leopard, Cheetah, and numbers of the smaller carnivora. Antelope include Eland, Sable, Roan, Liehtenstein's Hartebeest, Blue Wildebeest, Kudu, Defassa Waterbuck, Bushbuck, Reedbuck, Puku, Impala, Oribi, Common, Blue, and Yellow-backed Duikers, Klipspringer, and Sharpe's Stein- buck. There are Red Lechwe and Sitatunga in the Busango Swamp in the north, Hippopotamus in numbers in the Kafue and its larger tributaries. Zebra are common, Warthog and Bushpig everywhere. Birds are abundant. The park is uninhabited apart from certain small settle- ments, on a limited section of the Kafue River, belonging to the indigenous Africans under their tribal chief Kayingu. These people must be accorded their traditional local hunting rights, but such will affect only a small fraction of the whole vast wild area. -

Capture, Restraint and Transport Stress in Southern Chamois (Rupicapra Pyrenaica)
Capture,Capture, restraintrestraint andand transporttransport stressstress ininin SouthernSouthern chamoischamois ((RupicapraRupicapra pyrenaicapyrenaica)) ModulationModulation withwith acepromazineacepromazine andand evaluationevaluation usingusingusing physiologicalphysiologicalphysiological parametersparametersparameters JorgeJorgeJorge RamónRamónRamón LópezLópezLópez OlveraOlveraOlvera 200420042004 Capture, restraint and transport stress in Southern chamois (Rupicapra pyrenaica) Modulation with acepromazine and evaluation using physiological parameters Jorge Ramón López Olvera Bellaterra 2004 Esta tesis doctoral fue realizada gracias a la financiación de la Comisión Interministerial de Ciencia y Tecnología (proyecto CICYT AGF99- 0763-C02) y a una beca predoctoral de Formación de Investigadores de la Universidad Autónoma de Barcelona, y contó con el apoyo del Departament de Medi Ambient de la Generalitat de Catalunya. Los Doctores SANTIAGO LAVÍN GONZÁLEZ e IGNASI MARCO SÁNCHEZ, Catedrático de Universidad y Profesor Titular del Área de Conocimiento de Medicina y Cirugía Animal de la Facultad de Veterinaria de la Universidad Autónoma de Barcelona, respectivamente, CERTIFICAN: Que la memoria titulada ‘Capture, restraint and transport stress in Southern chamois (Rupicapra pyrenaica). Modulation with acepromazine and evaluation using physiological parameters’, presentada por el licenciado Don JORGE R. LÓPEZ OLVERA para la obtención del grado de Doctor en Veterinaria, se ha realizado bajo nuestra dirección y, considerándola satisfactoriamente -

Anaplasma Phagocytophilum and Babesia Species Of
pathogens Article Anaplasma phagocytophilum and Babesia Species of Sympatric Roe Deer (Capreolus capreolus), Fallow Deer (Dama dama), Sika Deer (Cervus nippon) and Red Deer (Cervus elaphus) in Germany Cornelia Silaghi 1,2,*, Julia Fröhlich 1, Hubert Reindl 3, Dietmar Hamel 4 and Steffen Rehbein 4 1 Institute of Comparative Tropical Medicine and Parasitology, Ludwig-Maximilians-Universität München, Leopoldstr. 5, 80802 Munich, Germany; [email protected] 2 Institute of Infectology, Friedrich-Loeffler-Institut, Südufer 10, 17493 Greifswald Insel Riems, Germany 3 Tierärztliche Fachpraxis für Kleintiere, Schießtrath 12, 92709 Moosbach, Germany; [email protected] 4 Boehringer Ingelheim Vetmedica GmbH, Kathrinenhof Research Center, Walchenseestr. 8-12, 83101 Rohrdorf, Germany; [email protected] (D.H.); steff[email protected] (S.R.) * Correspondence: cornelia.silaghi@fli.de; Tel.: +49-0-383-5171-172 Received: 15 October 2020; Accepted: 18 November 2020; Published: 20 November 2020 Abstract: (1) Background: Wild cervids play an important role in transmission cycles of tick-borne pathogens; however, investigations of tick-borne pathogens in sika deer in Germany are lacking. (2) Methods: Spleen tissue of 74 sympatric wild cervids (30 roe deer, 7 fallow deer, 22 sika deer, 15 red deer) and of 27 red deer from a farm from southeastern Germany were analyzed by molecular methods for the presence of Anaplasma phagocytophilum and Babesia species. (3) Results: Anaplasma phagocytophilum and Babesia DNA was demonstrated in 90.5% and 47.3% of the 74 combined wild cervids and 14.8% and 18.5% of the farmed deer, respectively. Twelve 16S rRNA variants of A. phagocytophilum were delineated. -

Dama Dama. by George A
MAMMALIAN SPECIES No. 317, pp. 1-8,3 figs. Dama dama. By George A. Feldharner, Kelly C. Farris-Renner, and Celeste M. Barker Published 27 December 1988 by The American Society of Mammalogists Dama dama (Linnaeus, 1758) Detailed descripti ons of the skull and dentition of European fallow deer are in Flerov (19 52), and Harrison (1968) described Persian Fallow Deer fallow deer. Cercus dum a Linnaeus, 1758 :67. T ype locality Sweden (introduced). Downloaded from https://academic.oup.com/mspecies/article/doi/10.2307/3504141/2600626 by guest on 29 September 2021 Ty pe spec ies of Duma Frisch. 1775 validated by plenary GENERAL CHARACTERS. Pelage coloration is the most powers. var iable of any spec ies of deer, with four main color varieties: white. Plat vccros plinii Zimmer ma n, 1780: 129. Renamin g of duma. men il, common (typical), and black (Chapm an and Chapm an. 1975). Cercus platvccros G. Cuvier , 1798:160. Renaming of dama. Int ermediat e pelage colors are cream. sandy , silver-grey, and sooty C NI'IIS mauricus F. Cuvier, 18 16:72. No localit y given. (Whitehead. 1972). Typical pelage is darker on the dorsal surface Ccr ru s (I)ama) m esopot amicus Brooke. 1875:26 4. Type localit y than the ventral sur face . ches t. and lower legs. A black dor sal stripe Khuzistan, Luristan (Persia). Ir an . ext end s from the na pe of the neck to the tip of the tail and around the upper edge of the white rump patch. T ypically. white spots are CONTEXT AND CONTENT. -

Engl South Africa Limpopo, Game Reserves & Brandberg 2017
South Africa 2017 - Limpopo , Game Reserve s & Brandberg - Office Germany: Office Austria: Ziegelstadel 1 · D-88316 Isny Europastrasse 1/1 · A-7540 Güssing Phone.: +49 (0) 75 62 / 914 54 - 14 Phone: +43 (0) 33 22 / 42 963 - 0 www.blaser-safaris.com Fax.: +43 (0) 33 22 / 42 963 - 59 [email protected] Hunt in South In South Africa, safaris can be conducted throughout the year; however the best time Africa : for a safari is between April & October. Some 40 different species of game can be hunted across the country, but each safari is individually planned to give you maximum enjoyment & satisfaction. The safari is suitable for the whole family, for non-hunters; there are a variety of inter- esting, photographic tours. All wildlife species listed in the price list are hunted on farmland / hunting areas around the lodge. LIMPOPO Limpopo The area of our partner Wayne Wagner Safaris is located in Hoedspruit Limpopo Pro v- ince and is situated between Phalaborwa, Gravelotte and Mica on the Olifants River. Accommodation To be sure that the safari meets with your specific requirements a questionnaire will be for Limpopo & completed before your arrival with your personal details, species to be hunted, food Games Reserves preferences and accommodation requirements. - 2 - Prices 201 7 5 DAY SOUTH AFRICAN PLAINS GAME CULLING PACKAGE The following package is a cull hunt for certain plains game species. This package includes 5 full days of hunting; you must plan on arriving one day prior to commencement of the safari. The hunt will take place in the bushveld region of the Limpopo Province near the town of Hoedspruit. -

The Effect of Species Associations on the Diversity and Coexistence of African Ungulates
The effect of species associations on the diversity and coexistence of African ungulates. By Nancy Barker For Professor Kolasa BIO306H1 – Tropical Ecology University of Toronto Wednesday, August 24th, 2005 Abstract: The effects of species associations on species diversity and coexistence were investigated in East Africa. The frequency and group sizes of African ungulates were observed and analyzed to determine for differences in species associations based on their density and distribution, as well as their associations with other species. Associations between species were determined to be nonrandom and seen to affect the demographics of associating herds. Such associations mirrored in other studies were shown to be the result of interspecific competition, habitat preferences and predation pressure which increases the potential for coexistence between species. This suggests a potentially important role in the regulation of species diversity by ecological dynamics in species rich communities. In the face of today’s biodiversity crisis, such understanding of species associations and how they are regulated may have huge implications for conservation. Introduction: known with the famous Darwin’s finches of the Galapagos Islands. However, there are many other Sympatric coexistence of organisms within a guilds with what seems to be extensive community poses several questions for ecologists. overlapping in their resources, such as the grazing High levels of species association occur with high herds in Africa which eat common and widely species packing, as is seen within the Selous game dispersed foods. Sinclair (Sinclair, 1979 as cited in reserve of Tanzania in east Africa. Sinclair (1985) Sinclair, 1985) has found that this seemingly notes that mixed herds are frequently seen in east extensive overlap among these herds have also Africa and Connor and Simberloff (1979) have undergone niche separation. -

Monitoring the Recovery of Wildlife in the Parque Nacional Da Gorongosa Through Aerial Surveys 2000
Monitoring the recovery of wildlife in the Parque Nacional da Gorongosa through aerial surveys 2000 - 2012 A preliminary analysis Dr Marc Stalmans July 2012 Summary • A total of 7 aerial wildlife surveys have been undertaken from 2000 to 2012 in the Gorongosa National Park. These have been sampling surveys covering 9 to 22% of the Park. • The data of these 6 helicopter and 1 fixed-wing (2004) survey have not yet been fully analysed. The data were combined in a single data base of 15 083 individual species occurrence records. These data were also incorporated in a Geographic Information System. • The results clearly indicate that there has been since 2000 a significant increase in wildlife numbers for most species. The vast majority of the wildlife is found in the central and southern part of the fertile Rift Valley. However, densities are much lower in the infertile miombo in the east and west as well as in the Rift Valley closer to human habitation. Where higher levels of protection from illegal hunting are maintained, such as in the Sanctuario, higher densities of wildlife are now recorded even though the recovery started from similar low levels as in other parts of the Park. • The current sampling design has significant shortcomings that make the estimate of overall population numbers problematic, especially for species with relatively low numbers and a clumped distribution (e.g. buffalo and elephant). This makes it difficult to evaluate the current populations against historical wildlife numbers. • The current design is not considered good enough for the quality of data required for ecological research and for the auditing of management performance. -

Evolutionary History of Red Deer with Special Reference to Islands
Evolutionary History of Red Deer with Special Reference to Islands Adrian Lister, Natural History Museum, London Mitochondrial DNA phylogeny of red/sika deer Meiri et al 2017 elaphus hanglu nippon canadensis Currently suggested taxonomy Lorenzini & Garofalo 2015, Meiri et al 2017, IUCN 2017 C. elaphus C. hanglu C. nippon C. canadensis Suggested region of origin and dispersal RITA LORENZINI and LUISA GAROFALO 2015 Earliest red deer fossils European early Middle Pleistocene Kashmir stag C. hanglu (0.9 Ma) ‘Cervus acoronatus’ Later… European coronate red deer E. Asian wapiti type (C. canadensis) (C. elaphus) from 400 ka fossils modern distribution historical distribution Meiri et al 2017 The dwarf deer of Jersey Belle Hougue Cave Age 120 ka (Last Interglacial) Lister 1989, 1995 The bones are a small form of red deer, Cervus elaphus Shoulder ht Body mass Mainland 1.25-1.30 m 200-250 kg Jersey 0.7 m 36 kg Three ways to get onto an island: 1. You are already there. Sea level rises and cuts off the island 2. You swim or raft across open sea 3. You are taken there by people large deer La Cotte, Jersey – large deer large deer 150ka: 100m contour 125ka: 10m contour 6,000 years of isolation in the Last Interglacial. Dwarf form lost when Jersey reconnected in last glaciation. Mediterranean islands: degree of endemicity, and subspecies/species status, depends on time of isolation Praemegaceros ‘Pseudodama’ Megaloceros Cervus Dama d Eucladoceros a c b a b c CORSICA/SARDINIA d MALTA SICILY CRETE Praemegaceros cazioti Cervus spp. & C. e. corsicanus Dama carburangelensis & C.