The Effect of Species Associations on the Diversity and Coexistence of African Ungulates
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
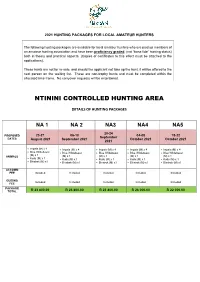
Ntinini Controlled Hunting Area
2021 HUNTING PACKAGES FOR LOCAL AMATEUR HUNTERS The following hunting packages are available for local amateur hunters who are paid up members of an amateur hunting association and have been proficiency graded, (not “bona fide” hunting status) both in theory and practical aspects. (Copies of certificates to this effect must be attached to the applications). These hunts are not for re-sale, and should the applicant not take up the hunt, it will be offered to the next person on the waiting list. These are non-trophy hunts and must be completed within the allocated time frame. No carryover requests will be entertained. NTININI CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES NA 1 NA 2 NA3 NA4 NA5 20-24 23-27 06-10 04-08 18-22 PROPOSED September DATES August 2021 September 2021 October 2021 October 2021 2021 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest (M) x 1 ANIMALS (M) x 1 (M) x 1 (M) x 1 (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 ACCOMM FEE Included Included Included Included Included GUIDING Included Included Included Included Included FEE PACKAGE TOTAL R 23 800.00 R 23 800.00 R 23 800.00 R 26 000.00 R 22 000.00 ITHALA CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES IGRA 1 IGRA 2 IGRA 3 IGRA 4 PROPOSED 11-16 02-07 16-21 06-11 DATES June 2021 July 2021 July 2021 August 2021 • Impala -

Annual Mass Drownings of the Serengeti Wildebeest Migration Influence Nutrient Cycling and Storage in the Mara River
Annual mass drownings of the Serengeti wildebeest migration influence nutrient cycling and storage in the Mara River Amanda L. Subaluskya,b,1, Christopher L. Duttona,b, Emma J. Rosib, and David M. Posta aDepartment of Ecology and Evolutionary Biology, Yale University, New Haven, CT 06511; and bCary Institute of Ecosystem Studies, Millbrook, NY 12545 Edited by James A. Estes, University of California, Santa Cruz, CA, and approved May 16, 2017 (received for review September 8, 2016) The annual migration of ∼1.2 million wildebeest (Connochaetes taur- hundreds to thousands of bison drowning in rivers of the western inus) through the Serengeti Mara Ecosystem is the largest remaining United States in the late 18th and early 19th centuries when overland migration in the world. One of the most iconic portions of large bison herds were still intact (16, 17). River basins that no their migration is crossing of the Mara River, during which thousands longer contain terrestrial migrations may have lost the annual drown annually. These mass drownings have been noted, but their input of resources from mass drownings, which may fundamen- frequency, size, and impact on aquatic ecosystems have not been tally alter how those river ecosystems function now compared quantified. Here, we estimate the frequency and size of mass drown- with the past. The more well-studied annual migrations of ings in the Mara River and model the fate of carcass nutrients through anadromous fishes and subsequent carcass inputs have been > the river ecosystem. Mass drownings ( 100 individuals) occurred in at shown to be important in rivers (18, 19), but equivalent exami- least 13 of the past 15 y; on average, 6,250 carcasses and 1,100 tons of nation of the influence of terrestrial migrations is lacking. -

Serengeti National Park
Serengeti • National Park A Guide Published by Tanzania National Parks Illustrated by Eliot Noyes ~~J /?ookH<~t:t;~ 2:J . /1.). lf31 SERENGETI NATIONAL PARK A Guide to your increased enjoyment As the Serengeti National Park is nearly as big as Kuwait or Northern Ireland no-one, in a single visit, can hope to see Introduction more than a small part of it. If time is limited a trip round The Serengeti National Park covers a very large area : the Seronera valley, with opportunities to see lion and leopard, 13,000 square kilometres of country stretching from the edge is probably the most enjoyable. of the Ngorongoro Conservation Unit in the south to the Kenya border in the north, and from the shores of Lake Victoria in the If more time is available journeys can be made farther afield, west to the Loliondo Game Controlled Area in the east. depending upon the season of the year and the whereabouts of The name "Serengeti" is derived from the Maasai language the wildlife. but has undergone various changes. In Maasai the name would be "Siringet" meaning "an extended area" but English has Visitors are welcome to get out of their cars in open areas, but replaced the i's with e's and Swahili has added a final i. should not do so near thick cover, as potentially dangerous For all its size, the Serengeti is not, of itself, a complete animals may be nearby. ecological unit, despite efforts of conservationists to make it so. Much of the wildlife· which inhabits the area moves freely across Please remember that travelling in the Park between the hours the Park boundaries at certain seasons of the year in search of 7 p.m. -

Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout Has in THIS ISSUE Dbeen Awarded the President’S Cup Letter from the President
DSC NEWSLETTER VOLUME 32,Camp ISSUE 5 TalkJUNE 2019 Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout has IN THIS ISSUE Dbeen awarded the President’s Cup Letter from the President .....................1 for her pending world record East African DSC Foundation .....................................2 Defassa Waterbuck. Awards Night Results ...........................4 DSC’s April Monthly Meeting brings Industry News ........................................8 members together to celebrate the annual Chapter News .........................................9 Trophy and Photo Award presentation. Capstick Award ....................................10 This year, there were over 150 entries for Dove Hunt ..............................................12 the Trophy Awards, spanning 22 countries Obituary ..................................................14 and almost 100 different species. Membership Drive ...............................14 As photos of all the entries played Kid Fish ....................................................16 during cocktail hour, the room was Wine Pairing Dinner ............................16 abuzz with stories of all the incredible Traveler’s Advisory ..............................17 adventures experienced – ibex in Spain, Hotel Block for Heritage ....................19 scenic helicopter rides over the Northwest Big Bore Shoot .....................................20 Territories, puku in Zambia. CIC International Conference ..........22 In determining the winners, the judges DSC Publications Update -

Serengeti: Nature's Living Laboratory Transcript
Serengeti: Nature’s Living Laboratory Transcript Short Film [crickets] [footsteps] [cymbal plays] [chime] [music plays] [TONY SINCLAIR:] I arrived as an undergraduate. This was the beginning of July of 1965. I got a lift down from Nairobi with the chief park warden. Next day, one of the drivers picked me up and took me out on a 3-day trip around the Serengeti to measure the rain gauges. And in that 3 days, I got to see the whole park, and I was blown away. [music plays] I of course grew up in East Africa, so I’d seen various parks, but there was nothing that came anywhere close to this place. Serengeti, I think, epitomizes Africa because it has everything, but grander, but louder, but smellier. [music plays] It’s just more of everything. [music plays] What struck me most was not just the huge numbers of antelopes, and the wildebeest in particular, but the diversity of habitats, from plains to mountains, forests and the hills, the rivers, and all the other species. The booming of the lions in the distance, the moaning of the hyenas. Why was the Serengeti the way it was? I realized I was going to spend the rest of my life looking at that. [NARRATOR:] Little did he know, but Tony had arrived in the Serengeti during a period of dramatic change. The transformation it would soon undergo would make this wilderness a living laboratory for understanding not only the Serengeti, but how ecosystems operate across the planet. This is the story of how the Serengeti showed us how nature works. -

Influence of Common Eland (Taurotragus Oryx) Meat Composition on Its Further Technological Processing
CZECH UNIVERSITY OF LIFE SCIENCES PRAGUE Faculty of Tropical AgriSciences Department of Animal Science and Food Processing Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing DISSERTATION THESIS Prague 2018 Author: Supervisor: Ing. et Ing. Petr Kolbábek prof. MVDr. Daniela Lukešová, CSc. Co-supervisors: Ing. Radim Kotrba, Ph.D. Ing. Ludmila Prokůpková, Ph.D. Declaration I hereby declare that I have done this thesis entitled “Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing” independently, all texts in this thesis are original, and all the sources have been quoted and acknowledged by means of complete references and according to Citation rules of the FTA. In Prague 5th October 2018 ………..………………… Acknowledgements I would like to express my deep gratitude to prof. MVDr. Daniela Lukešová CSc., Ing. Radim Kotrba, Ph.D. and Ing. Ludmila Prokůpková, Ph.D., and doc. Ing. Lenka Kouřimská, Ph.D., my research supervisors, for their patient guidance, enthusiastic encouragement and useful critiques of this research work. I am very gratefull to Ing. Petra Maxová and Ing. Eva Kůtová for their valuable help during the research. I am also gratefull to Mr. Petr Beluš, who works as a keeper of elands in Lány, Mrs. Blanka Dvořáková, technician in the laboratory of meat science. My deep acknowledgement belongs to Ing. Radek Stibor and Mr. Josef Hora, skilled butchers from the slaughterhouse in Prague – Uhříněves and to JUDr. Pavel Jirkovský, expert marksman, who shot the animals. I am very gratefull to the experts from the Natura Food Additives, joint-stock company and from the Alimpex-maso, Inc. -

A National Park in Northern Rhodesia 15
A National Park in Northern Rhodesia 15 A NATIONAL PARK IN NORTHERN RHODESIA By a Proclamation dated 20th April, 1950, the Governor of Northern Rhodesia has established a National Park, to be known as the Kafue National Park. This new park covers some 8,650 square miles roughly in the central Kafue basin, between latitudes 14° and 16° 40' S. It contains a wide range of country from the comparatively drv sandy lands of the south to the big rivers, swamps, and heavy timber of the northern section. The magnificent Kafue River dominates the whole central portion, adding scenic beauty to the attraction of wild life. The park contains representatives of most species of the fauna of Northern Rhodesia. Primates are represented by the Rhodesian Baboon, the Vervet Monkey, and the Greater and Lesser Night-Apes. There are Elephant and Black Rhinoceros, Buffalo, Lion, Leopard, Cheetah, and numbers of the smaller carnivora. Antelope include Eland, Sable, Roan, Liehtenstein's Hartebeest, Blue Wildebeest, Kudu, Defassa Waterbuck, Bushbuck, Reedbuck, Puku, Impala, Oribi, Common, Blue, and Yellow-backed Duikers, Klipspringer, and Sharpe's Stein- buck. There are Red Lechwe and Sitatunga in the Busango Swamp in the north, Hippopotamus in numbers in the Kafue and its larger tributaries. Zebra are common, Warthog and Bushpig everywhere. Birds are abundant. The park is uninhabited apart from certain small settle- ments, on a limited section of the Kafue River, belonging to the indigenous Africans under their tribal chief Kayingu. These people must be accorded their traditional local hunting rights, but such will affect only a small fraction of the whole vast wild area. -

Safari Drive Bingo
Safari Drive Bingo myfreebingocards.com Play Print off your bingo cards and start playing! If you can't get to a printer you can also play online - share this link with your friends: myfreebingocards.com/M/VrQeb and they can play on their mobiles or tablets. On the next page is a sheet for the bingo caller that contains of all the words that appear on the cards. To call the bingo you can cut the sheet up and pull the words out of a hat. Share Pin these bingo cards on Pinterest, share on Facebook, or post this link: myfreebingocards.com/S/VrQeb Edit and Create To add more words or make changes to this set of bingo cards go to myfreebingocards.com/E/VrQeb Go to myfreebingocards.com/bingo-card-generator to create a new set of bingo cards. Have Fun! If you have any feedback or suggestions about the bingo card generator, drop me an email on [email protected]. Bingo Caller's Card Cheetah Elephant Lion Buffalo Ostrich Birds Insects Zebra Giraffe Hyena Wild Dog Leopard Impala Hippo Gazelle Rhino Monkey Other Butterfly Antelope Wildebeest Mongoose Snake Jackal Warthog Snake Porcupine Squirrel myfreebingocards.com Safari Drive Bingo Safari Drive Bingo Mongoose Wildebeest Impala Zebra Porcupine Other Birds Giraffe FREE Porcupine Rhino Hippo Wild Dog Jackal Monkey Snake SPACE Monkey Elephant Squirrel Hyena Squirrel Insects Antelope Hyena FREE Insects Ostrich Birds Warthog Wildebeest Hippo Butterfly SPACE myfreebingocards.com myfreebingocards.com Safari Drive Bingo Safari Drive Bingo Hippo Other Snake Gazelle Birds Other Monkey Gazelle FREE -

Sustainable Use of Wildland Resources: Ecological, Economic and Social Interactions
Sustainable Use of Wildland Resources: Ecological, Economic and Social Interactions An Analysis of Illegal Hunting of Wildlife in Serengeti National Park, Tanzania Ken Campbell, Valerie Nelson and Martin Loibooki June 2001 Main Report This report should be cited as: Campbell, K. L. I., Nelson, V. and Loibooki, M. (2001). Sustainable use of wildland resources, ecological, economic and social interactions: An analysis of illegal hunting of wildlife in Serengeti National Park, Tanzania. Department for International Development (DFID) Animal Health Programme and Livestock Production Programmes, Final Technical Report, Project R7050. Natural Resources Institute (NRI), Chatham, Kent, UK. 56 pp. 2 Sustainable Use of Wildland Resources: Ecological, Economic and Social Interactions An Analysis of Illegal Hunting of wildlife in Serengeti National Park, Tanzania FINAL TECHNICAL REPORT, 2001 DFID Animal Health and Livestock Production Programmes, Project R7050 Ken Campbell1, Valerie Nelson2 Natural Resources Institute, University of Greenwich, Chatham, ME4 4TB, UK and Martin Loibooki Tanzania National Parks, P.O. Box 3134, Arusha, Tanzania Executive Summary A common problem for protected area managers is illegal or unsustainable extraction of natural resources. Similarly, lack of access to an often decreasing resource base may also be a problem for rural communities living adjacent to protected areas. In Tanzania, illegal hunting of both resident and migratory wildlife is a significant problem for the management of Serengeti National Park. Poaching has already reduced populations of resident wildlife, whilst over-harvesting of the migratory herbivores may ultimately threaten the integrity of the Serengeti ecosystem. Reduced wildlife populations may in turn undermine local livelihoods that depend partly on this resource. This project examined illegal hunting from the twin perspectives of conservation and the livelihoods of people surrounding the protected area. -

Human Pressure Threaten Swayne's Hartebeest to Point of Local
Research Article Volume 8:1,2020 Journal of Biodiversity and Endangered DOI: 10.24105/2332-2543.2020.8.239 Species ISSN: 2332-2543 Open Access Human Pressure Threaten Swayne’s Hartebeest to Point of Local Extinction from the Savannah Plains of Nech Sar National Park, South Rift Valley, Ethiopia Simon Shibru1*, Karen Vancampenhout2, Jozef Deckers2 and Herwig Leirs3 1Department of Biology, Arba Minch University, Arba Minch, Ethiopia 2Department of Earth and Environmental Sciences, Katholieke Universiteit Leuven, Celestijnenlaan 200E, B-3001 Leuven, Belgium 3Department of Biology, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerpen, Belgium Abstract We investigated the population size of the endemic and endangered Swayne’s Hartebeest (Alcelaphus buselaphus swaynei) in Nech Sar National Park from 2012 to 2014 and document the major threats why the species is on the verge of local extinction. The park was once known for its abundant density of Swayne’s Hartebeest. We used direct total count methods for the census. We administered semi-structured interviews and open-ended questionnaires with senior scouts who are a member of the local communities. Historical records were obtained to evaluate the population trends of the animals since 1974. The density of the animal decreased from 65 in 1974 to 1 individual per 100 km2 in 2014 with a decline of 98.5% in the past 40 years. The respondents agreed that the conservation status of the park was in its worst condition ever now with only 2 Swayne’s Hartebeest left, with a rapid decline from 4 individuals in 2012 and 12 individuals in 2009. Mainly hunting and habitat loss, but also unsuitable season of reproduction and shortage of forage as minor factors were identified as threats for the local extinction of the Swayne’s Hartebeests. -

Serengeti National Park Tanzania
SERENGETI NATIONAL PARK TANZANIA Twice a year ungulate herds of unrivalled size pour across the immense savanna plains of Serengeti on their annual migrations between grazing grounds. The river of wildebeests, zebras and gazelles, closely followed by predators are a sight from another age: one of the most impressive in the world. COUNTRY Tanzania NAME Serengeti National Park NATURAL WORLD HERITAGE SITE 1981: Inscribed on the World Heritage List under Natural Criteria vii and x. STATEMENT OF OUTSTANDING UNIVERSAL VALUE [pending] INTERNATIONAL DESIGNATION 1981: Serengeti-Ngorongoro recognised as a Biosphere Reserve under the UNESCO Man & Biosphere Programme (2,305,100 ha, 1,476,300 ha being in Serengeti National Park). IUCN MANAGEMENT CATEGORY II National Park BIOGEOGRAPHICAL PROVINCE East African Woodland/Savanna (3.05.04) GEOGRAPHICAL LOCATION In the far north of Tanzania 200 km west of Arusha, adjoining the Ngorongoro Conservation Area, between 1° 30' to 3° 20'S and 34° 00' to 35°15'E. DATES AND HISTORY OF ESTABLISHMENT 1929: Serengeti Game Reserve declared (228,600 ha) to preserve lions, previously seen as vermin; 1940: Declared a Protected Area; 1951: Serengeti National Park created, including Ngorongoro; boundaries were modified in 1959; 1981: Recognised as part of the Serengeti-Ngorongoro UNESCO Biosphere Reserve. LAND TENURE State, in Mara, Arusha and Shinyanga provinces. Administered by the Tanzanian National Parks Authority. AREA 1,476,300ha. It is contiguous in the southeast with Ngorongoro Conservation Area (809,440ha), in the southwest with Maswa Game Reserve (220,000ha), in the west with the Ikorongo-Grumeti Game Reserves (500,000ha), in the north with the Maasai-Mara National Reserve (151,000ha) in Kenya and in the northeast with the Loliondo Game Controlled Area (400,000ha). -

Connochaetes Gnou – Black Wildebeest
Connochaetes gnou – Black Wildebeest Blue Wildebeest (C. taurinus) (Grobler et al. 2005 and ongoing work at the University of the Free State and the National Zoological Gardens), which is most likely due to the historic bottlenecks experienced by C. gnou in the late 1800s. The evolution of a distinct southern endemic Black Wildebeest in the Pleistocene was associated with, and possibly driven by, a shift towards a more specialised kind of territorial breeding behaviour, which can only function in open habitat. Thus, the evolution of the Black Wildebeest was directly associated with the emergence of Highveld-type open grasslands in the central interior of South Africa (Ackermann et al. 2010). Andre Botha Assessment Rationale Regional Red List status (2016) Least Concern*† This is an endemic species occurring in open grasslands in the central interior of the assessment region. There are National Red List status (2004) Least Concern at least an estimated 16,260 individuals (counts Reasons for change No change conducted between 2012 and 2015) on protected areas across the Free State, Gauteng, North West, Northern Global Red List status (2008) Least Concern Cape, Eastern Cape, Mpumalanga and KwaZulu-Natal TOPS listing (NEMBA) (2007) Protected (KZN) provinces (mostly within the natural distribution range). This yields a total mature population size of 9,765– CITES listing None 11,382 (using a 60–70% mature population structure). This Endemic Yes is an underestimate as there are many more subpopulations on wildlife ranches for which comprehensive data are *Watch-list Threat †Conservation Dependent unavailable. Most subpopulations in protected areas are stable or increasing.