Download Complete Work
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

TREATISE ONLINE Number 48
TREATISE ONLINE Number 48 Part N, Revised, Volume 1, Chapter 31: Illustrated Glossary of the Bivalvia Joseph G. Carter, Peter J. Harries, Nikolaus Malchus, André F. Sartori, Laurie C. Anderson, Rüdiger Bieler, Arthur E. Bogan, Eugene V. Coan, John C. W. Cope, Simon M. Cragg, José R. García-March, Jørgen Hylleberg, Patricia Kelley, Karl Kleemann, Jiří Kříž, Christopher McRoberts, Paula M. Mikkelsen, John Pojeta, Jr., Peter W. Skelton, Ilya Tëmkin, Thomas Yancey, and Alexandra Zieritz 2012 Lawrence, Kansas, USA ISSN 2153-4012 (online) paleo.ku.edu/treatiseonline PART N, REVISED, VOLUME 1, CHAPTER 31: ILLUSTRATED GLOSSARY OF THE BIVALVIA JOSEPH G. CARTER,1 PETER J. HARRIES,2 NIKOLAUS MALCHUS,3 ANDRÉ F. SARTORI,4 LAURIE C. ANDERSON,5 RÜDIGER BIELER,6 ARTHUR E. BOGAN,7 EUGENE V. COAN,8 JOHN C. W. COPE,9 SIMON M. CRAgg,10 JOSÉ R. GARCÍA-MARCH,11 JØRGEN HYLLEBERG,12 PATRICIA KELLEY,13 KARL KLEEMAnn,14 JIřÍ KřÍž,15 CHRISTOPHER MCROBERTS,16 PAULA M. MIKKELSEN,17 JOHN POJETA, JR.,18 PETER W. SKELTON,19 ILYA TËMKIN,20 THOMAS YAncEY,21 and ALEXANDRA ZIERITZ22 [1University of North Carolina, Chapel Hill, USA, [email protected]; 2University of South Florida, Tampa, USA, [email protected], [email protected]; 3Institut Català de Paleontologia (ICP), Catalunya, Spain, [email protected], [email protected]; 4Field Museum of Natural History, Chicago, USA, [email protected]; 5South Dakota School of Mines and Technology, Rapid City, [email protected]; 6Field Museum of Natural History, Chicago, USA, [email protected]; 7North -

The Freshwater Bivalve Mollusca (Unionidae, Sphaeriidae, Corbiculidae) of the Savannah River Plant, South Carolina
SRQ-NERp·3 The Freshwater Bivalve Mollusca (Unionidae, Sphaeriidae, Corbiculidae) of the Savannah River Plant, South Carolina by Joseph C. Britton and Samuel L. H. Fuller A Publication of the Savannah River Plant National Environmental Research Park Program United States Department of Energy ...---------NOTICE ---------, This report was prepared as an account of work sponsored by the United States Government. Neither the United States nor the United States Depart mentof Energy.nor any of theircontractors, subcontractors,or theiremploy ees, makes any warranty. express or implied or assumes any legalliabilityor responsibilityfor the accuracy, completenessor usefulnessofanyinformation, apparatus, product or process disclosed, or represents that its use would not infringe privately owned rights. A PUBLICATION OF DOE'S SAVANNAH RIVER PLANT NATIONAL ENVIRONMENT RESEARCH PARK Copies may be obtained from NOVEMBER 1980 Savannah River Ecology Laboratory SRO-NERP-3 THE FRESHWATER BIVALVE MOLLUSCA (UNIONIDAE, SPHAERIIDAE, CORBICULIDAEj OF THE SAVANNAH RIVER PLANT, SOUTH CAROLINA by JOSEPH C. BRITTON Department of Biology Texas Christian University Fort Worth, Texas 76129 and SAMUEL L. H. FULLER Academy of Natural Sciences at Philadelphia Philadelphia, Pennsylvania Prepared Under the Auspices of The Savannah River Ecology Laboratory and Edited by Michael H. Smith and I. Lehr Brisbin, Jr. 1979 TABLE OF CONTENTS Page INTRODUCTION 1 STUDY AREA " 1 LIST OF BIVALVE MOLLUSKS AT THE SAVANNAH RIVER PLANT............................................ 1 ECOLOGICAL -

Guide to Estuarine and Inshore Bivalves of Virginia
W&M ScholarWorks Dissertations, Theses, and Masters Projects Theses, Dissertations, & Master Projects 1968 Guide to Estuarine and Inshore Bivalves of Virginia Donna DeMoranville Turgeon College of William and Mary - Virginia Institute of Marine Science Follow this and additional works at: https://scholarworks.wm.edu/etd Part of the Marine Biology Commons, and the Oceanography Commons Recommended Citation Turgeon, Donna DeMoranville, "Guide to Estuarine and Inshore Bivalves of Virginia" (1968). Dissertations, Theses, and Masters Projects. Paper 1539617402. https://dx.doi.org/doi:10.25773/v5-yph4-y570 This Thesis is brought to you for free and open access by the Theses, Dissertations, & Master Projects at W&M ScholarWorks. It has been accepted for inclusion in Dissertations, Theses, and Masters Projects by an authorized administrator of W&M ScholarWorks. For more information, please contact [email protected]. GUIDE TO ESTUARINE AND INSHORE BIVALVES OF VIRGINIA A Thesis Presented to The Faculty of the School of Marine Science The College of William and Mary in Virginia In Partial Fulfillment Of the Requirements for the Degree of Master of Arts LIBRARY o f the VIRGINIA INSTITUTE Of MARINE. SCIENCE. By Donna DeMoranville Turgeon 1968 APPROVAL SHEET This thesis is submitted in partial fulfillment of the requirements for the degree of Master of Arts jfitw-f. /JJ'/ 4/7/A.J Donna DeMoranville Turgeon Approved, August 1968 Marvin L. Wass, Ph.D. P °tj - D . dvnd.AJlLJ*^' Jay D. Andrews, Ph.D. 'VL d. John L. Wood, Ph.D. William J. Hargi Kenneth L. Webb, Ph.D. ACKNOWLEDGEMENTS The author wishes to express sincere gratitude to her major professor, Dr. -

Page 196 the Veliger, Vol. 32, No. 2
Page 196 The Veliger, Vol. 32, No. 2 Explanation of Figures 22 to 41 Figures 22-28. Adelodonax tectus sp. nov., from UCLA 3622. LACMIP 7843 from UCLA 6489, hypotype, right valve; Figure Figure 22: LACMIP 7825, holotype, left valve, xl. Figure 23: 31, exterior, xl; Figure 38, hinge, x2. Figure 32: LACMIP LACMIP 7827, paratype, hinge left valve, x3. Figure 24: 7841 from UCLA 3960, hypotype, hinge left valve, x2. Figure LACMIP 7826, paratype, hinge left valve, x3. Figure 25: LAC- 33: LACMIP 7840 from UCLA loc. 3958, hypotype, hinge right MIP 7829, paratype, right valve, x 1. Figure 26: LACMIP 7828, valve, x2. Figure 34: LACMIP 7842 from UCLA loc. 3960, paratype, right valve, pallial sinus, x2. Figure 27: LACMIP hypotype, "butterflied" valves, xl.5. Figure 35: LACMIP 7862 7830, paratype, hinge right valve, x 2. Figure 28: LACMIP 7831, from UCLA loc. 6489, hypotype, hinge left valve, x2. Figures paratype, hinge right valve, x 3. 36, 40: LACMIP 7837 from LACMIP 28629, hypotype, interior Figures 29-41. Adelodonax altus (Gabb, 1864). Figure 29: ANSP left valve, x 2; Figure 36, rock mold; Figure 40, latex pull. Figures 4557 from Martinez, Contra Costa Co., Calif., lectotype, x2. 37, 41: LACMIP 7839 from LACMIP loc. 28629, hypotype, Photo by Takeo Susuki. Figure 30: ANSP 71880 from Martinez, hinge right valve, x2; Figure 37, rock mold; Figure 41, latex Contra Costa Co., Calif., paralectotype, left valve showing trace pull. Figure 39: LACMIP 7838 from LACMIP loc. 28629, of pallial line, x2. Photo by Takeo Susuki. Figures 31, 38: hypotype, interior of left valve, latex pull, x 2. -

Manual to the Freshwater Mussels of MD
MMAANNUUAALL OOFF TTHHEE FFRREESSHHWWAATTEERR BBIIVVAALLVVEESS OOFF MMAARRYYLLAANNDD CHESAPEAKE BAY AND WATERSHED PROGRAMS MONITORING AND NON-TIDAL ASSESSMENT CBWP-MANTA- EA-96-03 MANUAL OF THE FRESHWATER BIVALVES OF MARYLAND Prepared By: Arthur Bogan1 and Matthew Ashton2 1North Carolina Museum of Natural Science 11 West Jones Street Raleigh, NC 27601 2 Maryland Department of Natural Resources 580 Taylor Avenue, C-2 Annapolis, Maryland 21401 Prepared For: Maryland Department of Natural Resources Resource Assessment Service Monitoring and Non-Tidal Assessment Division Aquatic Inventory and Monitoring Program 580 Taylor Avenue, C-2 Annapolis, Maryland 21401 February 2016 Table of Contents I. List of maps .................................................................................................................................... 1 Il. List of figures ................................................................................................................................. 1 III. Introduction ...................................................................................................................................... 3 IV. Acknowledgments ............................................................................................................................ 4 V. Figure of bivalve shell landmarks (fig. 1) .......................................................................................... 5 VI. Glossary of bivalve terms ................................................................................................................ -

<I>Fimbria Fimbriata</I>
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Morton, Brian, 1979. The biology and functional morphology of the coral- sand bivalve Fimbria fimbriata (Linnaeus 1758). Records of the Australian Museum 32(11): 389–420, including Malacological Workshop map. [31 December 1979]. doi:10.3853/j.0067-1975.32.1979.468 ISSN 0067-1975 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia 389 THE BIOLOGY AND FUNCTIONAL MORPHOLOGY OF THE CORAL-SAND BIVALVE FIMBRIA FIMBRIATA (Linnaeus 1758). BY BRIAN MORTON Department of Zoology, The University of Hong Kong SUMMARY Fimbria fimbriata Linnaeus 1758 is an infaunal inhabitant of coral sands in the Indo-Pacific. The structure and mineralogy of the shell (Taylor, Kennedy and Hall, 1973) confirms its taxonomic position as a member of the Lucinacea. Nicol (1950) erected (giving no reasons) a new family, taking its name (the Fimbriidae) from the genus. This study supports the view of Alien and Turner (1970) and Boss (1970) that Fimbria is closely related to the Lucinidae Fleming 1828 though a study of fossil fimbriids will have to be undertaken before the extreme view of Alien and Turner (1970) that Fimbria is a lucinid, can be validated. The Lucinidae and F. fimbriata possess the following features in common. 1. An enlarged anterior half of the shell with an antero-dorsal inhalant stream. 2. A single (inner) demibranch with type G ciliation (Atkins, 1937b). -

Bivalve Biology - Glossary
Bivalve Biology - Glossary Compiled by: Dale Leavitt Roger Williams University Bristol, RI A Aberrant: (L ab = from; erro = wonder) deviating from the usual type of its group; abnormal; wandering; straying; different Accessory plate: An extra, small, horny plate over the hinge area or siphons. Adapical: Toward shell apex along axis or slightly oblique to it. Adductor: (L ad = to; ducere = to lead) A muscle that draws a structure towards the medial line. The major muscles (usually two in number) of the bivalves, which are used to close the shell. Adductor scar: A small, circular impression on the inside of the valve marking the attachment point of an adductor muscle. Annulated: Marked with rings. Annulation or Annular ring: A growth increment in a tubular shell marked by regular constrictions (e.g., caecum). Anterior: (L ante = before) situated in front, in lower animals relatively nearer the head; At or towards the front or head end of a shell. Anterior extremity or margin: Front or head end of animal or shell. In gastropod shells it is the front or head end of the animal, i.e. the opposite end of the apex of the shell; in bivalves the anterior margin is on the opposite side of the ligament, i.e. where the foot protrudes. Apex, Apexes or Apices: (L apex = the tip, summit) the tip of the spire of a gastropod and generally consists of the embryonic shell. First-formed tip of the shell. The beginning or summit of the shell. The beginning or summit or the gastropod spire. The top or earliest formed part of shell-tip of the protoconch in univalves-the umbos, beaks or prodissoconch in bivalves. -

Bivalvia 101
Bivalvia 101 An Introduction to Marine Bivalves Paul ValentichValentich--ScottScott Curator of Malacology Santa Barbara Museum of Natural History Beautiful Bivalves of Southern California Class Bivalvia = two valves (shells) Clams,,,p,y mussels, scallops, oysters and their kin • Two part shell Bivalvia Clams, mussels, scallops, oysters and their kin • Two part shell • No head Bivalvia Clams, mussels, scallops, oysters and their kin • Two part shell • No head • Most are filter feeders Bivalvia Clams, mussels, scallops, oysters and their kin • Two part shell • No head • Most are filter feeders • Broadcast spawners Bivalvia Clams, mussels, scallops, oysters and their kin • Bivalves are great to EAT!!! Bivalve Orienteering Lookinggp from the top – Dorsal view Bival ve O ri ent eeri ng Posterior Anterior How do we know which end is which?? Bival ve O ri ent eeri ng Ligament Posterior Anterior Liggypament is usually posterior Bival ve O ri ent eeri ng BkBeaks Posterior Anterior Beaks often point anterior Bival ve O ri ent eeri ng Left valve Posterior Anterior Right valve Determining left from right Bival ve O ri ent eeri ng Left hand Left valve Anterior pointed away from you Right hand Right valve Anterior pointed Quick hint away from you Bivalve Orienteering Dorsal Posterior Anterior Ventral Looking from the side – Lateral view Bivalve Orienteering Dorsal Posterior Anterior Ventral Looking on the inside Bivalve Characters Umbo Beak Beak vs. Umbo Bivalve Characters –Sculpture– Sculpture Radial Sculpture Bivalve Characters –Sculpture– Sculpture -

Morphological Description of Cyrtopleura Costata (Bivalvia: Pholadidae) from Southern Brazil
ARTICLE Morphological description of Cyrtopleura costata (Bivalvia: Pholadidae) from southern Brazil Nicole Stakowian¹ & Luiz Ricardo L. Simone² ¹ Universidade Federal do Paraná (UFPR), Setor de Ciências Biológicas, Departamento de Zoologia (DZOO), Programa de Pós-Graduação em Zoologia. Curitiba, PR, Brasil. ORCID: http://orcid.org/0000-0002-3031-783X. E-mail: [email protected] ² Universidade de São Paulo (USP), Museu de Zoologia (MZUSP). São Paulo, SP, Brasil. ORCID: http://orcid.org/0000-0002-1397-9823. E-mail: [email protected] Abstract. The aim of the study is to describe in detail, for the first time, the internal and external anatomy of Cyrtopleura costata, which displays ellipsoid and elongated valves with beige periostracum, the anterior adductor muscle unites the valves in the pre- umbonal region, with abduction capacity in its dorsal half, sparing the ligament. Two accessory valves are identified: the mesoplax (calcified) located in the umbonal region; and the protoplax (corneus) above the anterior adductor muscle. Internally there is a pair of well-developed apophysis that supports the labial palps and the pedal muscles, and support part of the gills. The posterior half of mantle ventral edge is fused and richly muscular, working as auxiliary adductor muscle. The siphons are completely united with each other, the incurrent being larger than the excurrent. The foot is small (about ⅛ the size of the animal). The kidneys extend laterally on the dorsal surface, solid, presenting a brown/reddish color. The style sac is well developed and entirely detached from the adjacent intestine. The intestine has numerous loops and curves within the visceral mass. The fecal pellets are coin-shaped. -

Pages 875-887
lIo.1139. DEEP-WATER MOLLUSCA-VERRILL AND BUSH. 875 TQ'Wet'; its groove is narrow and inconspicuous. The hinge-margin itself is rather thin and bears very numerous, erect, V-shaped, acute teeth which number, in the large specimens, aboutsixteen or seventeen in the anterior series and about thirty-two or thirty·three in t~e poste· rior, including a number of small proximal ones; beneath the beaks there is a smooth, edentulous space, often a little thickened at the inner margin and projecting a little inward in the middle, and continuing inside the series of small proximal teeth on each side. In some cases this thickened border seems to arise anteriorly and to pass under the posterior seriea, as a slight fold; in ot,her cases it is continued directly from one series to the other. The posterior series is nearly straight and about twice as long as the anterior which is strongly curved and distally somewhat recedes from the thin dor8al margin. The .anterior end of the shell is rather short and evenly rounded; the posterior is about twice as long, a little wider owing to a slight veutral expansion, compressed and obtusely rounded or subtruneated at the margin, but without any distinct carination or angulation. The pallial sinus is very broad and deep, extending nearly to the middle of the shell. The inner surface is smooth, white or grayish white. The exterior is smooth, except for the delieate lines of growth, and covered with a thin, brilliantly iridescent epidermis which, in live specimens, is pale yellowish green, but in dead valves is pale straw color. -
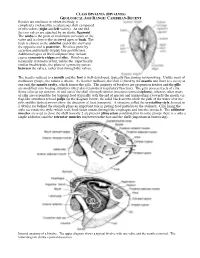
Bivales Are Molluscs in Which the Body Is Completely Enclosed by a Calcareous Shell Composed of Two Valves (Right and Left Valves)
CLASS BIVALVIA (BIVALVES) GEOLOGICAL AGE RANGE: CAMBRIAN-RECENT Bivales are molluscs in which the body is completely enclosed by a calcareous shell composed of two valves (right and left valves). At one end, the two valves are attached by an elastic ligament. The umbo is the point of maximum curvature of the valve and is close to the incurved apex or beak. The beak is closest to the anterior end of the shell and the opposite end is posterior. Bivalves grow by accretion and usually display fine growth lines. Additional types of shell sculpture may include coarse concentric ridges and ribs. Bivalves are bilaterally symmetrical but, unlike the superficially similar brachiopods, the plane of symmetry passes between the valves, rather than through the valves. The head is reduced to a mouth and the foot is well-developed, typically functioning in burrowing. Unlike most of molluscan groups, the radula is absent. As in other molluscs, the shell is lined by the mantle and there is a cavity at one end, the mantle cavity, which houses the gills. The majority of bivalves are suspension feeders and the gills are modified into feeding structures (they also retain their respiratory function). The gills possess tracts of cilia. Some cilia set up currents in and out of the shell (through tubular structures termed siphons), whereas other tracts of cilia are responsible for trapping food (typically with the aid of mucus) and transporting it towards the mouth via flap-like structures termed palps (in the diagram below, the solid black arrows show the path of the water over the gills and the dashed arrows show the direction of food transport). -

HOLE 317A, MANIHIKI PLATEAU Erle G
16. DEEP-SEA CRETACEOUS MACROFOSSILS: HOLE 317A, MANIHIKI PLATEAU Erle G. Kauffman, Department of Paleobiology, U.S. National Museum, Smithsonian Institution, Washington, D.C. INTRODUCTION Bivalves and other benthos are highly sensitive in- dicators of modern marine bottom environments, and Previous deep-sea cores penetrating Mesozoic strata thus of ancient sedimentary environments and their in- have failed to yield significant numbers of benthonic terpretation in deep-sea cores. In addition, molluscs are macrofossils, especially molluscs, other than fragments today (Hall, 1964), and in the past (e.g., Kauffman, of calcitic outer prismatic shell layers presumed, on the 1973a, b), primary organisms in the determination of basis of internal structure, to belong to the ubiquitous biogeographic units (realms, regions, provinces, sub- bivalve Family Inoceramidae. In part this paucity of provinces) and the study of their evolution. material has been attributed to unfavorable environ- Equally important is the biostratigraphic potential of ments of deep benthonic sediments. In part it has been molluscs that might be obtained from deep-sea cores; blamed on dissolution of shell material below the car- ammonites and bivalves are the most common Mesozoic bonate compensation depth, and especially of unstable molluscan elements in these samples and both are of aragonite which is an important component of many primary importance to refined zonation and regional mollusc shells. Aragonite compensation depth is about correlation. The mobile, widespread, and rapidly evolv- 3500 meters today; the major zone of dissolution lies ing ammonites are among the best biostratigraphic in- shallower, at a few hundred meters, where the rate of dicators known. Kauffman (1970, 1972, 1973a, b, 1975, aragonite solution is highest (Kennedy, 1969, p.