Genetic Resources of the Genus Cucumis and Their Morphological Description (English-Czech Version)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Cucurbitaceae), Hodgsonia Is the Hodgsoniinae C. Jeffrey, Subfamily
BLUMEA 46 (2001) 169-179 Taxonomy of Hodgsonia (Cucurbitaceae), with a note on the ovules and seeds W.J.J.O. de Wilde & B.E.E. Duyfjes Nationaal Herbarium Nederland, Universiteit Leiden branch, P.O. Box 9514, 2300 RA Leiden, The Netherlands Summary for Hodgsonia, ranging from NE India through S China to Java and Borneo, was a long time con- sidered as monotypic, but there are two (and possibly three) species, demarcated at the Isthmus of Kra in S Thailand. The ‘seeds’ should be condition known few, woody regarded as pyrenes, a not elsewhere in the family Cucurbitaceae. words Flora of SE Key . Asia, Cucurbitaceae, Hodgsonia,placentation, pyrenes. Introduction Hodgsonia is the only genus in the subtribe Hodgsoniinae C. Jeffrey, tribe Trichosan- theae C. Jeffrey, subfamily Cucurbitoideae(Jeffrey, 1962). The subtribe is definedby having 12 ovules, in 6 collateralpairs, the seeds connate in pairs, one of a pair usually smallerand with an abortive embryo. The connate seeds form large seed-like pyrenes. Within the tribe, Hodgsonia is also palynologically distinct (Khunwasi, 1998). Hodgsonia contains two species as presented here, both with edible oil in their seeds There has been confusion the (Burkill, 1935; Hu, 1964). on delimitation, name, and range of the two species, to such an extent that in practically all literaturethe con- cept ofthe used species names is contaminated.The history of the genusand the des- ignation of its various species names has been related by Hu (1964). The two currently accepted species appear to occupy different, adjacent areas in SE Asia. Theirdistribu- tions are distinct and meet somewhere in the area of S Tenasserim in S Myanmar or line the Isthmus of Kra in S Thailand, but the precise demarcation is as yet unknown because of insufficient collectionsfrom that area. -

The Heirloom Gardener's Seed-Saving Primer Seed Saving Is Fun and Interesting
The Heirloom Gardener's Seed-Saving Primer Seed saving is fun and interesting. It tells the story of human survival, creativity, and community life. Once you learn the basics of saving seeds you can even breed your own variety of crop! Share your interesting seeds and stories with other gardeners and farmers while helping to prevent heirloom varieties from going extinct forever. Contact The Foodshed Project to find out about local seed saving events! 1. Food “as a system”...........................................................................................................................5 2. Why are heirloom seeds important?.................................................................................................6 3. How are plants grouped and named?...............................................................................................8 4. Why is pollination important?... ......................................................................................................11 5. What is a monoecious or a dioecious plant?....................................................................................12 6. How do you know if a plant will cross-breed?.................................................................................14 7. What types of seeds are easiest to save?........................................................................................18 8. What about harvesting and storing seeds?.....................................................................................20 9. What do I need to know -

The Genus Trichosanthes L. (Cucurbitaceae) in Thailand
THAI FOR. BULL. (BOT.) 32: 76–109. 2004. The genus Trichosanthes L. (Cucurbitaceae) in Thailand BRIGITTA E.E. DUYFJES*& KANCHANA PRUESAPAN** ABSTRACT. Trichosanthes (Cucurbitaceae) in Thailand comprises 17 species, seven of which have been described as new here: T. dolichosperma Duyfjes & Pruesapan, T. erosa Duyfjes & Pruesapan, T. inthanonensis Duyfjes & Pruesapan, T. kostermansii Duyfjes & Pruesapan, T. pallida Duyfjes & Pruesapan, T. phonsenae Duyfjes & Pruesapan, and T. siamensis Duyfjes & Pruesapan. Two new subspecific entities have been described: T. pubera Blume subsp. rubriflos (Cayla) Duyfjes & Pruesapan var. fissisepala Duyfjes & Pruesapan, and T. tricuspidata Lour. subsp. javanica Duyfjes & Pruesapan. A key to taxa, descriptions with distributional and ecological data and illustrations are presented. INTRODUCTION The results of the present revision of Trichosanthes will form part of the forthcoming treatment of the family Cucurbitaceae for the Flora of Thailand. Trichosanthes is an Asian genus, extending eastward to Australia. Within the Cucurbitaceae of Thailand, as well as for the whole of Southeast Asia, it is the largest genus, with 17 species in Thailand, and more than 100 species in all. It is a difficult genus, not because the species are unclear, but the herbaria materials are generally insufficient because the species are dioecious, the fragile corollas which bloom at night are difficult to collect, to preserve and to study, and the fruiting collections (fruits are quite often collected) at first sight show little relation to the flowering specimens. Trichosanthes is known as medicinal, and has recently received comparatively much taxonomic attention in China (Yueh & Cheng, 1974, 1980). The genus has been revised for India by Chakravarty (1959), for Cambodia, Laos and Vietnam by Keraudren (1975), and for the Malesian area by Rugayah & De Wilde (1997, 1999) and Rugayah (1999). -

An Ethanobotanical Investigation Of
Journal of Medicinal Plants Studies 2017; 5(3): 250-254 ISSN (E): 2320-3862 ISSN (P): 2394-0530 An ethanobotanical investigation of cucurbitaceae NAAS Rating 2017: 3.53 JMPS 2017; 5(3): 250-254 from South India: A review © 2017 JMPS Received: 05-03-2017 Accepted: 06-04-2017 Thammaihraj Shanthi Avinash and Vittal Ravishankar Rai Thammaihraj Shanthi Avinash Department of Studies in Abstract Microbiology, University of Cucurbitaceae crops are cash crops grown as vegetables. The family cucurbitaceae well known medicinal Mysore, Manasagangothri, uses the whole plant parts including roots, leaves, fruits and seeds have been extensively studied for their Mysore, Karnataka, India. pharmacological activity. A herb is a plant that is valued for flavor, scent, or other qualities. From ancient Vittal Ravishankar Rai days to now a day, plant parts used in cooking, as medicines as they were potential and treatment of Department of Studies in several diseases and disorders. Main behind of that is medicinal plants is not having any side effects. Microbiology, University of Fruits of which are widely used in ayurveda and other folk medicines traditionally used for its Mysore, Manasagangothri, cardioprotective, cardiotonic, general tonic, diuretic, aphrodisiac, antidote to certain poisons and scorpion Mysore, Karnataka, India. strings, alternative purgative, cooling effects. It cures pain, ulcers and fever and used for pectoral cough, asthma and other bronchial disorders-especially syrup prepared from the tender fruits. The fruit is reported to contain the triterepeniode cucurbitacins B, D, G, H and 22-deoxy cucurbitacin. This is an attempt to compile an up-to-date and comprehensive review of that covers its traditional and folk medicinal uses, phytochemistry and pharmacology. -

Trichosanthes Dioica Roxb.: an Overview
PHCOG REV. REVIEW ARTICLE Trichosanthes dioica Roxb.: An overview Nitin Kumar, Satyendra Singh, Manvi, Rajiv Gupta Department of Pharmacognosy, Faculty of Pharmacy, Babu Banarasi Das National Institute of Technology and Management, Dr. Akhilesh Das Nagar, Faizabad Road, Lucknow, Uttar Pradesh, India Submitted: 01-08-2010 Revised: 05-08-2011 Published: 08-05-2012 ABSTRACT Trichosanthes, a genus of family Cucurbitaceae, is an annual or perennial herb distributed in tropical Asia and Australia. Pointed gourd (Trichosanthes dioica Roxb.) is known by a common name of parwal and is cultivated mainly as a vegetable. Juice of leaves of T. dioica is used as tonic, febrifuge, in edema, alopecia, and in subacute cases of enlargement of liver. In Charaka Samhita, leaves and fruits find mention for treating alcoholism and jaundice. A lot of pharmacological work has been scientifically carried out on various parts of T. dioica, but some other traditionally important therapeutical uses are also remaining to proof till now scientifically. According to Ayurveda, leaves of the plant are used as antipyretic, diuretic, cardiotonic, laxative, antiulcer, etc. The various chemical constituents present in T. dioica are vitamin A, vitamin C, tannins, saponins, alkaloids, mixture of noval peptides, proteins tetra and pentacyclic triterpenes, etc. Key words: Cucurbitacin, diabetes, hepatoprotective, Trichosanthes dioica INRODUCTION parmal, patol, and potala in different parts of India and Bangladesh and is one of the important vegetables of this region.[3] The fruits The plants in Cucurbitaceae family is composed of about 110 and leaves are the edible parts of the plant which are cooked in genera and 640 species. The most important genera are Cucurbita, various ways either alone or in combination with other vegetables Cucumis, Ecballium, Citrullus, Luffa, Bryonia, Momordica, Trichosanthes, or meats.[4] etc (more than 30 species).[1] Juice of leaves of T. -

Effects of Cucumis Metuliferus (Cucurbitaceae) Fruits on Enzymes and Haematological Parameters in Albino Rats
African Journal of Biotechnology Vol. 6 (22) pp. 2515-2518, 19 November, 2007 Available online at http://www.academicjournals.org/AJB ISSN 1684–5315 © 2007 Academic Journals Full Length Research Paper Effects of Cucumis metuliferus (Cucurbitaceae) fruits on enzymes and haematological parameters in albino rats Noel N. Wannang*, Nanloh S. Jimam, Simeon Omale, Maxwell L. P. Dapar, Steven S. Gyang and John C. Aguiyi Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmaceutical Sciences, University of Jos, Nigeria. Accepted 24 October, 2007 The effects of the powdered fruits of Cucumis metuliferus on enzymes and haematological indices were evaluated in adult albino rats. The study revealed a significant (P<0.05) dose-dependent decrease in white blood cells (WBC) count. 500 mg/kg body weight of the powdered fruit produced a significant (P<0.05) decrease in red blood cells (RBC), and an increase in platelet and haemoglobin (Hb), while there was an insignificant (P>0.05) decrease in clotting and bleeding time. 1000 mg/kg produced significant (P<0.05) increase in RBC, platelets, Hb and packed cell volume (PCV) values, and an insignificant (P>0.05) decrease in clotting and bleeding time. The biochemical parameters evaluation showed that 500 - 1000 mg/kg of the powdered fruit of the plant produced a dose-dependent significant (P<0.05) increase in the levels of serum alkaline phosphatase (ALP), aspartate aminotransferase (AST), alanine aminotransferase (ALT), Blood urea nitrogen (BUN) and Total protein. This result showed that Cucumis metuliferus produced alterations in the haematological and biochemical indices evaluated. Keywords: Cucumis metuliferus, serum, enzymes, haematological. -

Notes on Cyclanthera Brachystachya (Ser.) Cog.: Morphology, Ecology, and Its New Distribution
pISSN 2302-1616, eISSN 2580-2909 Vol 7, No. 1, Juni 2019, hal 54-57 Available online http://journal.uin-alauddin.ac.id/index.php/biogenesis DOI https://doi.org/10.24252/bio.v7i1.6239 Notes on Cyclanthera brachystachya (Ser.) Cog.: Morphology, Ecology, and Its New Distribution MUHAMMAD EFENDI1*, RUGAYAH2 1Cibodas Botanic Gardens - LIPI Jl. Raya Kebun Raya Cibodas, Cianjur, West Java, Indonesia. 43253. 2Herbarium Bogoriense, Botany Division, Research Center for Biology - LIPI Jl. Raya Jakarta - Bogor Km. 46 Cibinong, Bogor, Indonesia. 16911. *Email: [email protected] Received 11 October 2018; Received in revised form 10 December 2018; Accepted 19 June 2019; Available online 30 June 2019 ABSTRACT Cyclanthera brachystachya (Cucurbitaceae) is an exotic species in Indonesia that originated from America. This species was reported running wild in several places in Java for decade ago. However, it is limited information to support this statement. This needs to be done, because C. brachystachya have potentially to increase the genetic resources of cucumber family in Indonesia. The methods used include morphological and ecological observation, including their distribution from their naturalized habitat in Indonesia. Morphology of this species were described based on living material from Cibodas Botanical Garden area. Morphologically, it is not different with other morphology description on flora of Java and other literatures. In their natural habitat, they grow on open areas with high humidity levels and variously substrate. This species spreads in the mountain of West Java, from Mt. Gede Pangrango to Mt. Manglayang Bandung and Garut. A new distribution in Sumatera added outside range of this species in their origin area in Indonesia. -

Evolution of Angiosperm Pollen. 7. Nitrogen-Fixing Clade1
Evolution of Angiosperm Pollen. 7. Nitrogen-Fixing Clade1 Authors: Jiang, Wei, He, Hua-Jie, Lu, Lu, Burgess, Kevin S., Wang, Hong, et. al. Source: Annals of the Missouri Botanical Garden, 104(2) : 171-229 Published By: Missouri Botanical Garden Press URL: https://doi.org/10.3417/2019337 BioOne Complete (complete.BioOne.org) is a full-text database of 200 subscribed and open-access titles in the biological, ecological, and environmental sciences published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Complete website, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/terms-of-use. Usage of BioOne Complete content is strictly limited to personal, educational, and non - commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Downloaded From: https://bioone.org/journals/Annals-of-the-Missouri-Botanical-Garden on 01 Apr 2020 Terms of Use: https://bioone.org/terms-of-use Access provided by Kunming Institute of Botany, CAS Volume 104 Annals Number 2 of the R 2019 Missouri Botanical Garden EVOLUTION OF ANGIOSPERM Wei Jiang,2,3,7 Hua-Jie He,4,7 Lu Lu,2,5 POLLEN. 7. NITROGEN-FIXING Kevin S. Burgess,6 Hong Wang,2* and 2,4 CLADE1 De-Zhu Li * ABSTRACT Nitrogen-fixing symbiosis in root nodules is known in only 10 families, which are distributed among a clade of four orders and delimited as the nitrogen-fixing clade. -

Iran Has the Second Largest Economy (After Saudi Arabia) and Population (After Egypt)
COUNTRY FACT SHEET ON FOOD AND AGRICULTURE POLICY TRENDS | September 2014 Socio-economic context and role of agriculture In the Near East and North Africa region, Iran has the second largest economy (after Saudi Arabia) and population (after Egypt). Iran ranks second in the world in natural gas reserves and third in oil reserves.1 The agriculture and rural sector share in the GDP has declined in the last twenty years and yet is the source of income for more than 15 million people in rural areas.2 One quarter of the rural population is landless and of those who own land, one third are smallholders. Those within this segment of the population often fall below or just within the poverty line and face high underemployment rates. IRAN Some of the main development challenges at the national level are the harsh conditions of the physical environment and low productivity of small-holder farmers. Food security challenges include lack of self-sufficiency in major staple crops and inadequate access to food in terms of quantity of daily energy intake. The Iranian Government has adopted a comprehensive strategy envisioning market-based reforms as reflected in the 20-year Vision document and Iran’s fifth Five-Year Development Plan (FYDP 2011–15). The Government envisioned a large privatization programme in its 2010-2015 five-year plan, aiming to privatize 20 percent of state-owned enterprises (SOEs) each year. Moreover, Iran’s 2012 Doing Business ranking is in the bottom tiers of the Middle East and North Africa (MENA) region, at 144th overall. Only Algeria, Iraq, and Djibouti rank lower among MENA countries. -
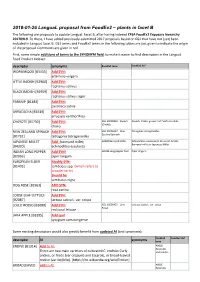
2018-01-26 Langual Proposal from Foodex2 – Plants in Facet B
2018-01-26 LanguaL proposal from FoodEx2 – plants in facet B The following are proposals to update LanguaL Facet B, after having indexed EFSA FoodEx2 Exposure hierarchy 20170919. To these, I have added previously-submitted 2017 proposals based on GS1 that have not (yet) been included in LanguaL facet B. GS1 terms and FoodEx2 terms in the following tables are just given to indicate the origin of the proposal. Comments are given in red. First, some simple additions of terms to the SYNONYM field, to make it easier to find descriptors in the LanguaL Food Product Indexer: descriptor synonyms FoodEx2 term FoodEx2 def WORMWOOD [B3433] Add SYN: artemisia vulgaris LITTLE RADISH [B2960] Add SYN: raphanus sativus BLACK RADISH [B2959] Add SYN: raphanus sativus niger PARSNIP [B1483] Add SYN: pastinaca sativa ARRACACHA [B3439] Add SYN: arracacia xanthorrhiza CHAYOTE [B1730] Add SYN: GS1 10006356 - Squash Squash, Choko, grown from Sechium edule (Choko) choko NEW ZEALAND SPINACH Add SYN: GS1 10006427 - New- Tetragonia tetragonoides Zealand Spinach [B1732] tetragonia tetragonoides JAPANESE MILLET Add : barnyard millet; A000Z Barnyard millet Echinochloa esculenta (A. Braun) H. Scholz, Barnyard millet or Japanese Millet. [B4320] echinochloa esculenta INDIAN LONG PEPPER Add SYN! A019B Long pepper fruit Piper longum [B2956] piper longum EUROPEAN ELDER Modify SYN: [B1403] sambucus spp. (which refers to broader term) Should be sambucus nigra DOG ROSE [B2961] ADD SYN: rosa canina LOOSE LEAF LETTUCE Add SYN: [B2087] lactusa sativa L. var. crispa LOLLO ROSSO [B2088] Add SYN: GS1 10006425 - Lollo Lactuca sativa L. var. crispa Rosso red coral lettuce JAVA APPLE [B3395] Add syn! syzygium samarangense Some existing descriptors would also greatly benefit from updated AI (and synonyms): FoodEx2 FoodEx2 def descriptor AI synonyms term ENDIVE [B1314] Add to AI: A00LD Escaroles There are two main varieties of cultivated C. -

High Tunnel Melon and Watermelon Production
High Tunnel Melon and Watermelon Production University of Missouri Extension M173 Contents Author Botany 1 Lewis W. Jett, Division of Plant Sciences, University of Missouri-Columbia Cultivar selection 3 Editorial staff Transplant production 4 MU Extension and Agricultural Information Planting in the high tunnel 5 Dale Langford, editor Dennis Murphy, illustrator Row covers 6 On the World Wide Web Soil management and fertilization 6 Find this and other MU Extension publications on the Irrigation 7 Web at http://muextension.missouri.edu Pollination 7 Photographs Pruning 8 Except where noted, photographs are by Lewis W. Jett. Trellising 8 Harvest and yield 9 Marketing 10 Pest management 10 Useful references 14 Melon and watermelon seed sources 15 Sources of high tunnels (hoophouses) 16 For further information, address questions to College of Dr. Lewis W. Jett Agriculture Extension State Vegetable Crops Specialist Food and Natural Division of Plant Sciences Resources University of Missouri Columbia, MO 65211 Copyright 2006 by the University of Missouri Board of Curators E-mail: [email protected] College of Agriculture, Food and Natural Resources High Tunnel Melon and Watermelon Production igh tunnels are low-cost, passive, melo has several botanical subgroups (Table 1). solar greenhouses that use no fossil In the United States, reticulatus and inodorus are Hfuels for heating or venting (Figure commercially grown, while the remaining groups 1). High tunnels can provide many benefits to are grown for niche or local markets. horticulture crop producers: The cantaloupe fruit that most Americans • High tunnels are used to lengthen the are familiar with is not actually a true cantaloupe. -

CUCUMBER LEMONGRASS MARTINI Ingredients: Directions: • 2 Oz Tito’S Vodka Muddle Cucumber in Shaker
COEUR D’COCKTAILS, JUSTIN SCHORZMAN CUCUMBER LEMONGRASS MARTINI Ingredients: Directions: • 2 oz Tito’s Vodka Muddle cucumber in shaker. Then add vodka, • 1 oz fresh lemon juice lemon juice, lemongrass simple syrup, and • 1.5 oz lemongrass lemonade in shaker. Fill shaker with ice, cover, simple syrup and shake vigorously until outside of shaker is • 1 oz fresh lemonade very cold, about 10 seconds. • 4 slices of cucumber Strain cocktail into a martini glass. Top with roasted lemon peel. COEUR D’COCKTAILS, JUSTIN SCHORZMAN CAPRESE BLOODY MARY Ingredients: Directions: • 1.5 oz Tito’s Vodka Lightly muddle four basil leaves in shaker. Then • 5 leaves fresh basil add vodka and house made bloody mary mix. • 5 oz house-made Fill shaker with ice, cover, and shake for about bloody mary mix 10 seconds. Rim pint glass with sea salt. Fill • 2 cherry tomatoes pint glass with ice. Strain bloody mary into pint • fresh mozzarella glass. Garnish with fresh mozzarella, cherry tomatoes, and basil leaf. COEUR D’COCKTAILS, JUSTIN SCHORZMAN GINGER PEAR SALTY DOG Ingredients: Directions: • 1.5 oz Tito’s Vodka Muddle pear wedges in shaker. Then add • 0.5 oz agave nectar vodka, agave nectar, and grapefruit juice. • 4 wedges of ripe Grate ginger root into shaker. Fill shaker with Asian pear ice, cover, and shake vigorously until outside • 4 oz fresh ruby red of shaker is very cold, about 10 seconds. grapefruit juice Rim Collins glass with Himalayan sea salt, fill • 1/4 tsp fresh ginger with ice. Strain cocktail into glass. Garnish with pear wedge..