The Influence of Neighborhood Landscape Characteristics on Native Bird Communities: Implications for Increasing Biodiversity in Our Yards
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Report on the 2003 Parks Levy Investment Objective 1: Restore
A Report on the 2003 Parks Levy Investment In November 2002, Portland voters approved a five-year Parks Levy to begin in July 2003. Levy dollars restored budget cuts made in FY 2002-03 as well as major services and improvements outlined in the Parks 2020 Vision plan adopted by City Council in July 2001. In order to fulfill our obligation to the voters, we identified four key objectives. This report highlights what we have accomplished to date. Objective 1: Restore $2.2 million in cuts made in 2002/03 budget The 2003 Parks Levy restored cuts that were made to balance the FY 2002-03 General Fund budget. These cuts included the closure of some recreational facilities, the discontinuation and reduction of some community partnerships that provide recreational opportunities for youth, and reductions in maintenance of parks and facilities. Below is a detailed list of services restored through levy dollars. A. Restore programming at six community schools. SUN Community Schools support healthy social and cross-cultural development of all participants, teach and model values of respect and inclusion of all people, and help reduce social disparities and inequities. Currently, over 50% of students enrolled in the program are children of color. 2003/04 projects/services 2004/05 projects/services Proposed projects/services 2005/06 Hired and trained full-time Site Coordinators Total attendance at new sites (Summer Continue to develop programming to serve for 6 new PP&R SUN Community Schools: 2004-Spring 2005): 85,159 the needs of each school’s community and Arleta, Beaumont, Centennial, Clarendon, increase participation in these programs. -
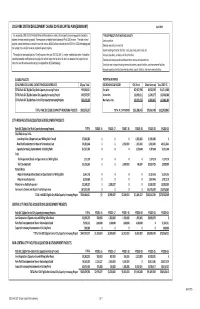
2015 DRAFT Park SDC Capital Plan 150412.Xlsx
2015 PARK SYSTEM DEVELOPMENT CHARGE 20‐YEAR CAPITAL PLAN (SUMMARY) April 2015 As required by ORS 223.309 Portland Parks and Recreation maintains a list of capacity increasing projects intended to TYPES OF PROJECTS THAT INCREASE CAPACITY: address the need created by growth. These projects are eligible to be funding with Park SDC revenue . The total value of Land acquisition projects summarized below exceeds the potential revenue of $552 million estimated by the 2015 Park SDC Methodology and Develop new parks on new land the funding from non-SDC revenue targeted for growth projects. Expand existing recreation facilities, trails, play areas, picnic areas, etc The project list and capital plan is a "living" document that, per ORS 223.309 (2), maybe modified at anytime. It should be Increase playability, durability and life of facilities noted that potential modifications to the project list will not impact the fee since the fee is not based on the project list, but Develop and improve parks to withstand more intense and extended use rather the level of service established by the adopted Park SDC Methodology. Construct new or expand existing community centers, aquatic facilities, and maintenance facilities Increase capacity of existing community centers, aquatic facilities, and maintenance facilities ELIGIBLE PROJECTS POTENTIAL REVENUE TOTAL PARK SDC ELIGIBLE CAPACITY INCREASING PROJECTS 20‐year Total SDC REVENUE CATEGORY SDC Funds Other Revenue Total 2015‐35 TOTAL Park SDC Eligible City‐Wide Capacity Increasing Projects 566,640,621 City‐Wide -

About East Portland Neighborhoods Vol
EAST PORTLAND NEIGHBORHOOD ASSOCIATION NEWS October 2009 News about East Portland Neighborhoods vol. 14 issue 4 Your NEIGHBORHOOD ASSOCIATIONS Argay pg.pg. pg. pg.5 pg.6 pg. Neighborhood Association 33 4 6 12 Centennial Community Association All about East Portland Glenfair Neighborhood Association Neighborhood Association News … Hazelwood The East Portland in outer East Portland that events, graffiti cleanups, and tribution with positive, far- Neighborhood Association Neighborhood Association make up the EPNO coalition tree plantings. reaching results. News (EPNAN) isn’t a news- (our alliance individual neigh- As you look through our The volunteers of the East Lents paper in the traditional sense. borhoods) – know more paper and see how your Portland Neighbors Inc. Neighborhood Association It wasn’t created to compete about this sanctioned system neighbors are making a real Newspaper Committee thank with community, city or of neighborhood organiza- difference in their neighbor- you for taking a few minutes Mill Park national news outlets – nei- tions, recognized by City gov- hood, perhaps you’ll be to discover more about what Neighborhood Association ther in content nor for adver- ernment. encouraged by their efforts. your neighbors are doing, tisers. So, the stories and photos Then, possibly you’ll decide and how you can help outer Parkrose Heights EPNAN is the way the East you see on the pages inside to take as little as one hour a East Portland be an even Association of Neighbors Portland Neighborhood are about volunteers and month to participate in your nicer place to live when we Organization (EPNO) reach- organizations that are work- neighborhood association work together. -

ORDINANCE NO. 187150 As Amended
ORDINANCE NO. 187150 As Amended Accept Park System Development Charge Methodology Update Report for implementation, and amend the applicable sections of City Code (Ordinance; amend Code Chapter 17.13) The City of Portland ordains: Section 1. The Council finds: 1. Ordinance No. 172614, passed by the Council on August 19, 1998 authorized establishment of a Parks and Recreation System Development Charge(SDC) and created a new City Code Chapter 17.13. 2. In October 1998 the City established a Parks SDC program. City Code required that the program be updated every two years to ensure that program goals were being met. An update was implemented on July 1, 2005 pursuant to Ordinance No. 179008, as amended. The required update reviewed the Parks SDC Program to determine that sufficient money will be available to fund capacity-increasing facilities identified by the Parks SDC-CIP; to determine whether the adopted and indexed SDC rate has kept pace with inflation; to determine whether the Parks SDC-CIP should be modified; and to ensure that SDC receipts will not over-fund such facilities. 3. Ordinance No 175774, passed by the Council on July 12, 2001 adopted The Parks 2020 Vision. This report highlighted significant challenges confronting the City in regards to shoring up our ailing park facilities, eliminating inequity in underserved neighborhoods, and providing a stable source of funding to address not just our existing shortfalls, but to also meet the needs created by new development. The Park SDC is the most significant revenue opportunity available to Parks to address growth. It is imperative that this opportunity is maximized to recover reasonable costs from new development. -

Sub-Area: Southeast
PARKS 2020 VISION OUTHEAST Distinctive Features Studio in the Laurelhurst Park annex is a satellite of the Montavilla Community Center. I Aquatic facilities include Sellwood, Mt. Scott, Description: The Southeast sub-area (see map at the Buckman, Montavilla and Creston. end of this section) contains many of the city's older, I established neighborhoods. This area is a patchwork of The Community Music Center is in this sub-area. older, mainly single-family neighborhoods divided by I The Southeast sub-area has three Community linear commercial corridors. The Central Eastside Schools and 45 school sites. Industrial District, which borders the east bank of the I There are lighted baseball stadiums at Willamette, separates some residential neighborhoods Westmoreland and Lents Parks. from the river. Resources and Facilities: Southeast has 898 acres Population – Current and Future: The Southeast of parkland, ranking third in total amount of park sub-area ranks first in population with 154,000 and acreage. Most parks are developed, well distributed, is projected to grow to 157,830 by 2020, an increase in good condition, and can accommodate a range of of 2%. recreational uses. I Southeast has the City’s largest combined acreage DISTRIBUTION OF SUBAREA ACRES BY PARK TYPE of neighborhood and community parks. I Southeast has a variety of habitat parks, including Oaks Bottom Wildlife Refuge, Tideman Johnson Park, and Johnson Creek Park that are popular sites for hiking, birding, walking, and general recreation use. I This sub-area includes part of the I-205 Bike Trail and about 4.6 miles of the Springwater Corridor, a 195-acre 16.5 mile-long regional trailway that includes many natural resources. -

Download PDF File 2019-20
2019–20 YEAR 5 PARKS BOND EXECUTIVE SUMMARY YEAR 5 Dear Portlanders, We are happy to report that 46 of the 52 Bond projects have been completed, with the remaining six projects underway. Your investment has been used wisely. Year 5 of the Bond started out as planned: • In July 2019, Commissioner Fish cut the ribbon on the completely overhauled Peninsula Pool. • In October 2019, the community gathered to celebrate a more accessible playground at Glenhaven Park. • Construction wrapped up on the installation of new play pieces and drainage repairs at over 30 parks. Sadly, 2020 started off with the loss of our colleague and beloved Parks Commissioner, Nick Fish. And then COVID-19 hit. With some adaptations, Bond projects stayed on track. Construction began on a new playground for Creston Park, and we completed a new playground at Verdell Burdine Rutherford Park, the first Portland park to be named solely after a Black woman. The public health crisis was followed by a groundswell of action for racial justice. Now, our parks and open spaces are even more precious than ever, serving as shared public spaces to exercise our bodies, our minds, and our voices. While this Bond could only tackle the most critical maintenance needs, it has given us all a glimpse of what we can achieve together. Let’s continue to create a more sustainable and more equitable future for our city and our parks. Stay safe, stay healthy, and stay hopeful. Sincerely, Commissioner Amanda Fritz Portland Parks & Recreation Director Adena Long 1 PARKS BOND EXECUTIVE SUMMARY YEAR 5 46Projects completed Glenhaven Park playground opening celebration Projects6 underway Peninsula Pool opening celebration Current3 projects ahead of or on schedule Marshall Park bridge construction 2 PARKS BOND EXECUTIVE SUMMARY YEAR 5 NORTH Acquisitions at Cathedral, Open Meadow. -

2016 Park System Development Charge 20-Year Capital Plan (Summary)
187770 Exhibit A 2016 PARK SYSTEM DEVELOPMENT CHARGE 20-YEAR CAPITAL PLAN (SUMMARY) As required by ORS 223.309 Portland Parks and Recreation maintains a list of capacity increasing projects intended to address the need created by growth. These projects are eligible to be funded with Park SDC revenue. The total value of projects summarized below exceeds the potential revenue of $552 million estimated by the 2015 Park SDC Methodology and the funding from non-SDC revenue targeted for growth projects. The project list and capital plan is a "living" document that, per ORS 223.309 (2), may be modified at any time. Changes to this list will not affect the SDC rates, unless the Council holds a public hearing and authorizes the changes, as provided in ORS 223.309(2). TYPES OF PROJECTS THAT INCREASE CAPACITY: Land acquisition Develop new parks on new land Expand existing recreation facilities, trails, play areas, picnic areas, etc Increase playability, durability and life of facilities Natural area restoration Develop and improve parks to withstand more intense and extended use Construct new or expand existing community centers, aquatic facilities, and maintenance facilities Increase capacity of existing community centers, aquatic facilities, and maintenance facilities SDC Zone Program Site Project Name % Growth Years 1 - 5 Years 6 - 10 Years 11 -10 Total 20 Years Total * Growth % Central City Acquisitions Central City Unidentified Central City Acquisitions 100% $ 5,000,000 $ 5,000,000 $ 5,000,000 Central City Acquisition Placeholder Downtown 100% -

District Background
DRAFT EAST PORTLAND DISTRICT PROFILE July 13, 2005 DRAFT Acknowledgements City of Portland Bureau of Planning Gil Kelley, Planning Director Joe Zehnder, Principal Planner District Profile Staff Barry Manning, Senior Planner, District Liaison Denver Igarta, Community Service Aide Harper Kalin, Community Service Aide GIS/Graphics Gary Odenthal Carmen Piekarski Ralph Sanders For more information please contact: City of Portland Bureau of Planning 1900 SW Fourth Avenue, Suite 4100 Portland, Oregon 97201 Phone: 503-823-7700 Fax: 503-823-7800 http://www.portlandonline.com/planning DRAFT East Portland District Profile Table of Contents Introduction Area Description ……………………………………………………………………. 1 Demographics Data ….……………………………………………………………. 5 Organization ……………………………………………………………………. 6 Neighborhood Facilities & Services ………………..…………………………… 9 Urban Renewal Areas ……………………………………………………………. 16 Land Use ……………………………………………………………………………. 19 Environment ……………………………………………………………………. 27 Development Activity ……………………………………………………………. 30 Economic Development ……………………………………………………………. 35 Transportation ……………………………………………………………………. 38 Existing Plans and Policies ……………………………………………………. 48 Major Capital Improvement Projects ………….…………………………………. 51 District-wide Issues .……………………………………………………………. 52 Potential Projects ………..………………………..…………………………………. 53 DRAFT DRAFT Introduction In 2004 the Bureau of Planning launched the District Liaison Program which assigns a City Planner to each of Portland’s designated liaison districts. Each planner acts as the Bureau’s primary contact -

DATE: March 26, 2008 TO: Board Of
DATE: March 26, 2008 TO: Board of Commissioners FROM: Bruce A. Warner, Executive Director SUBJECT: Report Number 08-36 Update: Lents Town Center Plan Amendment Study – Final Report & Recommendations EXECUTIVE SUMMARY BOARD ACTION REQUESTED None ─ information only. SUMMARY In April 2007 the Portland Development Commission (PDC) Board of Commissioners (Board) directed staff to conduct a study of the Lents Town Center Urban Renewal Area (URA). In July 2007, the Lents Town Center Urban Renewal Advisory Committee (URAC) appointed a subcommittee that was charged with developing policy recommendations for URAC consideration and PDC Board review and approval. The subcommittee was responsible for making recommendations and answering three fundamental questions: (1) Should the URA boundary be expanded and where? (2) Should the maximum indebtedness be increased to complete existing projects in the URA Plan, continue funding existing programs, and consider new projects identified by the community? (3) Should the current expiration date (2015) be extended? At the March 26, 2008, Board meeting, the Lents Team will provide an update and present a final report that documents the technical, policy alternatives, and public involvement process, as well as the URAC recommendations. Staff will seek comments on the core questions above and request authorization to move forward with the presentation of the URA Plan Amendment & Report on May 14, 2008. BACKGROUND The Lents Plan Amendment Study process included an extensive public outreach process with community stakeholders, including the URAC, neighborhood and business associations, and property owners and local businesses, and the general public. The subcommittee, which was responsible for the development of recommendations, held five evening meetings open to the public from August to December 2007. -

1 Getting to Know Lents: a Thematic Atlas Of
Getting to Know LentsGetting to Know Lents: A Thematic Atlas of HealthyA Thematic EatingAtlas of Healthy and Eating Active and Active Living Living Getting to Know Lents A Thematic Atlas of Healthy Eating and Active Living Getting to Know Lents: A Thematic Atlas of Healthy Eating and Active Living Getting to Know Lents A Thematic Atlas of Healthy Eating and Active Living Acknowledgements This project was made possible by Portland State University spring and summer capstone students 2008 in conjunction with Community Health Partnership: Oregon’s Public Health Institute. Spring 2008 Capstone Summer 2008 Capstone Community Partners Allison Adcox Ben Blessing 1000 Friends of Oregon Oregon Coalition for Marina Carter Preston Brookfield Active Living by Design Promoting Physical Allen Davis Valerie DePan American Heart Association Activity Jonathan Gray Sarah Egan Bureau of Planning Portland Development Devon Kelley Rory Hammock Coalition for a Livable Commission Lyn Kirby Brandon Jones Future Portland Office of Yu-Ching Liu Nick Jones Community Health Transportation Meg Merrick Troy Kenyon Partnership Portland Parks and Nick Nicholson James Kerridge Growing Gardens Recreation Steven Zach Owen Meg Merrick Kelly Elementary SUN Portland State University Blake Shepard Derrak Richard Program Portland/Multnomah Food Simon Skiles Michael Russell Lents Food Group Council Blia Xiong Hiroko Segawa Lents Neighborhood Robert Wood Johnson Blair Whiteman Association Foundation Marshall High School Wattles Boys and Girls Club Northwest Health Zenger Farm Foundation Getting to Know Lents A Thematic Atlas of Healthy Eating and Active Living Contents Mission Statement Acknowledgements ........................................................................ 4 Created through a lens of healthy eating and active Background on Community Health ............................................. 6 Why Place (Lents) Matters: Building a Movement for living, this atlas is intended to describe the historical Healthy Communities .................................................................. -

Comprehensive Annual Financial Report Exhibit a February 13, 2008 Page 1 of 177
Board Resolution 6558 - Comprehensive Annual Financial Report Exhibit A February 13, 2008 Page 1 of 177 Investing in Portland’s Future Exhibit A Page 2 of 177 On the Cover: Board Resolution 6558 - Comprehensive Annual Financial Report February 13, 2008 UNDER THE AUTUMN MOON FESTIVAL 2006 Culminating a planning process that began December 2005, the Portland Develop- ment Commisison (PDC) spearheaded an effort in September 2006 to celebrate the $5.3 million in streetscape improvements in Old Town/Chinatown (OTCT). More than 35,000 people attended the two and a half-day public event, called “Under the Autumn Moon.” The festivities kicked off Friday evening , September 29, with a ribbon cutting on NW Davis Street. A full day of activities and events began Saturday, September 30, with a parade (including grand marshal Mayor Tom Potter), two non-stop music stages, food booths, cooking demonstrations, a glob- al bazaar, tours of the district, an outdoor movie and a fireworks display. Events continued on Sunday, October 1, when the Chinese Classical Garden was open for free all day. Restaurants and businesses in OTCT participated by inviting crowds in to sample foods and wares. Many of the restaurants were packed throughout the event. Many people attending were from the Chinese community, some of whom had not been in Old Town/Chinatown for years. We hope this will be the first of many such events on the new festival streets and that those who attended the Under the Autumn Moon Festival will return often. The vision for the streetscape improvements was to strengthen the identity of the historic district, foster cultural and economic diversity, and promote a vibrant pe- destrian environment for commercial, retail and residential uses. -

Phase II Environmental Site Assessment Boys and Girls Club
Phase II Environmental Site Assessment Boys and Girls Club 9330 SE Harold Street Portland, Oregon October 1, 2010 Prepared for Portland Development Commission Portland, Oregon 333 SW 5th Avenue, Suite 700 Portland, OR 97204 (503) 542-1080 TABLE OF CONTENTS Page INTRODUCTION 1 BACKGROUND 1 WORK PERFORMED 2 ANALYTICAL RESULTS 4 CONCLUSIONS AND RECOMMENDATIONS 5 USE OF THIS REPORT 6 REFERENCES 8 FIGURES Figure Title 1 Vicinity Map 2 Anomalies and Subsurface Investigation Locations TABLES Table Title 1 Summary of Geophysical Anomaly Exploration 2 Test Pit Analytical Results 3 Soil Boring Analytical Results APPENDICES Appendix Title A Photograph Log B Boring Logs and Geotechnical Hole Reports C Laboratory Analytical Reports 10/1/10 I:\PROJECTS\1095\006\020\FILERM\R\FINAL PHASE II ESA\PHASE II ESA_RPT_FINAL.DOCX LANDAU ASSOCIATES ii INTRODUCTION This report summarizes the findings of a focused Phase II Environmental Site Assessment (ESA) conducted for the Lents Boys and Girls Club of Portland located at 9330 SE Harold Street in Portland, Oregon (subject property, Figure 1). The focused Phase II ESA was conducted by Landau Associates for the Portland Development Commission (PDC), based on the statement of work outlined in our scope of work dated July 28, 2010. This report summarizes the field activities conducted during the Phase II ESA and discusses the investigation findings and results. Conclusions based on the findings and recommendations for further action, as appropriate, are also included. BACKGROUND The Phase I ESA performed by Landau Associates (Landau Associates 2010) revealed evidence of two recognized environmental conditions in connection with the subject property: • Between 1935 and 1970, a gasoline/service station was located on the northeast corner of SE Harold Street and SE 92nd Avenue, directly across SE Harold Street from the subject property.