St. John and Cinnamon Bay
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Appendix A: Consultation and Coordination
APPENDIX A: CONSULTATION AND COORDINATION Virgin Islands National Park July 2013 Caneel Bay Resort Lease This page intentionally left blank Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-1 Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-2 Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-3 Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-4 Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-5 Virgin Islands National Park July 2013 Caneel Bay Resort Lease A-6 APPENDIX B: PUBLIC INVOLVEMENT Virgin Islands National Park July 2013 Caneel Bay Resort Lease This page intentionally left blank Virgin Islands National Park July 2013 Caneel Bay Resort Lease B-1 Virgin Islands National Park July 2013 Caneel Bay Resort Lease B-2 Virgin Islands National Park July 2013 Caneel Bay Resort Lease B-3 APPENDIX C: VEGETATION AND WILDLIFE ASSESSMENTS Virgin Islands National Park July 2013 Caneel Bay Resort Lease VEGETATION AND WILDLIFE ASSESSMENTS FOR THE CANEEL BAY RESORT LEASE ENVIRONMENTAL ASSESSMENT AT VIRGIN ISLANDS NATIONAL PARK ST. JOHN, U.S. VIRGIN ISLANDS Prepared for: National Park Service Southeast Regional Office Atlanta, Georgia March 2013 TABLE OF CONTENTS Page LIST OF FIGURES ...................................................................................................................... ii LIST OF TABLES ........................................................................................................................ ii LIST OF ATTACHMENTS ...................................................................................................... -

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

National List of Vascular Plant Species That Occur in Wetlands 1996
National List of Vascular Plant Species that Occur in Wetlands: 1996 National Summary Indicator by Region and Subregion Scientific Name/ North North Central South Inter- National Subregion Northeast Southeast Central Plains Plains Plains Southwest mountain Northwest California Alaska Caribbean Hawaii Indicator Range Abies amabilis (Dougl. ex Loud.) Dougl. ex Forbes FACU FACU UPL UPL,FACU Abies balsamea (L.) P. Mill. FAC FACW FAC,FACW Abies concolor (Gord. & Glend.) Lindl. ex Hildebr. NI NI NI NI NI UPL UPL Abies fraseri (Pursh) Poir. FACU FACU FACU Abies grandis (Dougl. ex D. Don) Lindl. FACU-* NI FACU-* Abies lasiocarpa (Hook.) Nutt. NI NI FACU+ FACU- FACU FAC UPL UPL,FAC Abies magnifica A. Murr. NI UPL NI FACU UPL,FACU Abildgaardia ovata (Burm. f.) Kral FACW+ FAC+ FAC+,FACW+ Abutilon theophrasti Medik. UPL FACU- FACU- UPL UPL UPL UPL UPL NI NI UPL,FACU- Acacia choriophylla Benth. FAC* FAC* Acacia farnesiana (L.) Willd. FACU NI NI* NI NI FACU Acacia greggii Gray UPL UPL FACU FACU UPL,FACU Acacia macracantha Humb. & Bonpl. ex Willd. NI FAC FAC Acacia minuta ssp. minuta (M.E. Jones) Beauchamp FACU FACU Acaena exigua Gray OBL OBL Acalypha bisetosa Bertol. ex Spreng. FACW FACW Acalypha virginica L. FACU- FACU- FAC- FACU- FACU- FACU* FACU-,FAC- Acalypha virginica var. rhomboidea (Raf.) Cooperrider FACU- FAC- FACU FACU- FACU- FACU* FACU-,FAC- Acanthocereus tetragonus (L.) Humm. FAC* NI NI FAC* Acanthomintha ilicifolia (Gray) Gray FAC* FAC* Acanthus ebracteatus Vahl OBL OBL Acer circinatum Pursh FAC- FAC NI FAC-,FAC Acer glabrum Torr. FAC FAC FAC FACU FACU* FAC FACU FACU*,FAC Acer grandidentatum Nutt. -

El Toro Wilderness, Luquillo Experimental Forest, Puerto Rico
El Toro Wilderness, Luquillo Experimental Forest, Puerto Rico Peter L. Weaver Abstract—The El Toro Wilderness, designated by Congress in 2005, man and his own works dominate the landscape, is hereby occupies about 36 percent of the 11,300 ha Luquillo Experimental recognized as an area where the earth and its community of Forest (LEF) in northeastern Puerto Rico. It is the only tropical life are untrammeled by man, where man himself is a visitor forest in the wilderness system managed by the U.S. Department who does not remain” (U.S. Department of Agriculture, For- of Agriculture. El Toro extends from 370 to 1,074 m in elevation, est Service 1983, pages 177-178). Wilderness areas provide and is occupied by four forest types found in the mountainous numerous advantages to society—ecological services such Caribbean: lower montane rain forest, montane rain forest, palm as clean air and water, natural habitat for flora and fauna, brake, and dwarf forest. The LEF, a Biosphere Reserve since 1976, scenic beauty, and solitude—and direct economic benefits to contains 225 tree species, 45 of them endemic to Puerto Rico, and surrounding communities through recreation and tourism 23 to the LEF alone; 150 species of ferns; 79 species of orchids; 11 (Dombeck 1999). native bats; 101 birds, 12 of them endemic to the island; 19 native In 1999, a new wilderness agenda was proposed that speci- reptiles, 8 endemic; 14 native amphibians, 8 endemic; and 6 na- fied six goals for the U.S. Forest Service (Dombeck 1999): tive fish species. Most of these species occupy the wilderness. -

Introduction to the Geography, Geology, Climate and Flora Habitats of Culebra Culebra Flora & Fauna Digital Database and Indexes Flora of Culebra, Puerto Rico
Introduction to the Geography, Geology, Climate and Flora Habitats of Culebra, Puerto Rico by: Manuel H. Dubón A Fundación Mi Terruño Flora Series Publication 2015 Edition Fundación Mi Terruño [FMT] – Culebra, Puerto Rico Introduction to the Geography, Geology, Climate and Flora Habitats of Culebra Culebra Flora & Fauna Digital Database and Indexes Flora of Culebra, Puerto Rico © Manuel H. Dubón Introduction The experience of appreciating the subtropical Flora and Fauna of Culebra affords visitors and residents a unique opportunity to enjoy a very special personal experience of both a visual and spiritual dimensions. Nature is a God given gift to be enjoyed today, shared, and conserved for our future generations. The FMT Culebra Flora and Fauna Digital Photographic Databases are published by Fundación Mi Terruño, Inc. (FMT) with special permission and license from its author and amateur photographer, Manuel H. Dubón. The database presents and describes the flora found within the land site of a proposed sustainable resort-residential development of advance design designated as Villa Mi Terruño (VMT). It has been expanded to cover general flora of Culebra. FMT publishes this photographic website to offer Culebra residents, our island visitors, guest from Puerto Rico or afar and website visitors a visual and learning experience as they enjoy the Flora of Culebra in its entire splendor. It will hopefully enrich the visitors and residents life experience as they appreciate and better understand their subtropical environment in the Caribbean island of Culebra. The Flora Digital Database will also allow website visitors, students and professional and amateur naturalist to do research and enjoy a specialized photographic database of the dry subtropical Flora of Culebra with ample references. -

Virgin Islands National Park Geologic Resources Inventory Report
National Park Service U.S. Department of the Interior Natural Resource Program Center Virgin Islands National Park Geologic Resources Inventory Report Natural Resource Report NPS/NRPC/GRD/NRR—2010/226 THIS PAGE: Underwater ecosystems including coral reefs are a primary natural resource at Virgin Islands National Park. National Park Service photograph. ON THE COVER: This view of Trunk Bay shows the steep slopes characteristic of Virgin Islands Na- tional Park. National Park Service photo- graph courtesy Rafe Boulon (Virgin Islands National Park). Virgin Islands National Park Geologic Resources Inventory Report Natural Resource Report NPS/NRPC/GRD/NRR—2010/226 Geologic Resources Division Natural Resource Program Center P.O. Box 25287 Denver, Colorado 80225 July 2010 U.S. Department of the Interior National Park Service Natural Resource Program Center Fort Collins, Colorado The National Park Service, Natural Resource Program Center publishes a range of reports that address natural resource topics of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Report Series is used to disseminate high-priority, current natural resource management information with managerial application. The series targets a general, diverse audience, and may contain NPS policy considerations or address sensitive issues of management applicability. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner. This report received informal peer review by subject-matter experts who were not directly involved in the collection, analysis, or reporting of the data. -

Fall/Winter 2016
A Palm Issue Seychelles & Madagascar page 3 US Virgin Islands page 4 Completing Our US Collection page 5 Montgomery Botanical Center Established 1959 Board of Directors David Manz, Esq., President Charles P. Sacher, Esq., Vice President Karl Smiley, M.D., Vice President Walter D. Haynes, Esq., Sec./Treasurer Charles S. Sacher, Esq., Asst. Treasurer Nicholas D. Kelly L. Patrick Kelly Peter A. Manz F r o m t h e Stephen D. Pearson, Esq. Juanita Popenoe, Ph.D. Executive Director Mark Smiley Executive Director Dear Friends, M. Patrick Griffith, Ph.D., M.B.A. am excited to send you this Palm Issue of our newsletter! As Research Fellows I promised in our previous issue, here we focus on Montgomery’s Angélica Cibrián Jaramillo, Ph.D. work with palms, and how that work helps advance the field of John Dowe, Ph.D. botanical science. William Hahn, Ph.D. Damon P. Little, Ph.D. At the center of our palm studies is Dr. Larry Noblick. Since Cristina Lopez-Gallego, Ph.D. 1994, Larry’s work for Montgomery has taken him around the globe, Mónica Moraes R., Ph.D. researching, collecting and conserving palms – over 2 decades of Fred Stauffer, Ph.D. building our collections. Larry’s first expedition to Madagascar in Alberto S. Taylor B., Ph.D. 1995 brought seed which are now stately Bismarckia, Borassus and Irene Terry, Ph.D. Hyphaene anchoring the landscape of our longest vistas. Thus, I am Barry Tomlinson, Ph.D. glad he shares his return to Madagascar, along with his work in the Seychelles, on the facing page. -

Biosíntesi, Distribució, Acumulació I Funció De La Vitamina E En Llavors: Mecanismes De Control
Biosíntesi, distribució, acumulació i funció de la vitamina E en llavors: mecanismes de control Laura Siles Suárez Aquesta tesi doctoral està subjecta a la llicència Reconeixement- NoComercial – SenseObraDerivada 3.0. Espanya de Creative Commons. Esta tesis doctoral está sujeta a la licencia Reconocimiento - NoComercial – SinObraDerivada 3.0. España de Creative Commons. This doctoral thesis is licensed under the Creative Commons Attribution-NonCommercial- NoDerivs 3.0. Spain License. Barcelona, febrer de 2017 Biosíntesi, distribució, acumulació i funció de la vitamina E en llavors: mecanismes de control Memòria presentada per Laura Siles Suarez per a optar al grau de Doctora per la Universitat de Barcelona. Aquest treball s’emmarca dins el programa de doctorat de BIOLOGIA VEGETAL del Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (BEECA) de la Facultat de Biologia de la Universitat de Barcelona. El present treball ha estat realitzat al Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals de la Facultat de Biologia (BEECA) de la Universitat de Barcelona sota la direcció de la Dra. Leonor Alegre Batlle i el Dr. Sergi Munné Bosch. Doctoranda: Directora i Codirector de Tesi: Tutora de Tesi: Laura Siles Suarez Dra. Leonor Alegre Batlle Dra. Leonor Alegre Batlle Dr. Sergi Munné Bosch “Mira profundamente en la naturaleza y entonces comprenderás todo mejor”- Albert Einstein. “La creación de mil bosques está en una bellota”-Ralph Waldo Emerson. A mi familia, por apoyarme siempre, y a mis bichejos peludos Índex ÍNDEX AGRAÏMENTS i ABREVIATURES v INTRODUCCIÓ GENERAL 1 Vitamina E 3 1.1.Descobriment i estudi 3 1.2.Estructura química i classes 3 Distribució de la vitamina E 5 Biosíntesi de vitamina E 6 3.1. -

Bay Native Plant Garden
WELCOME TO THE FISH BAY NATIVE PLANT GARDEN Tyre Palm This garden shows the types of plants and trees that grow best and naturally in Fish Bay. Before you excavate, use this garden area as a guide to what could grow on your land or may already be there. Tag trees on your building site for saving and only excavate the area covering the footprint of the home you are building. This garden is in greenspace owned by the Island Resources Foundation which permitted this education use. The Estate Fish Bay Owners’ Association was awarded a grant from the VI Department of Agriculture, Urban & Community Forestry Program to fund this project. Members of the community volunteered many hours to prepare the site, plant, and install the garden path and Eleanor Gibney, St John’s own botanist, guided us and was instrumental in creating this brochure. Thank you to all. Bay rum - Pimenta racemosa This beautiful native tree is the source of aromatic oil that is mixed with alcohol or rum to make the cologne or lotion known as bay rum which was extremely popular around the world in the period between 1880 and 1940. St. John was a major producer of bay oil. Locally, leaves are also used in home remedies and cooking. A medium sized evergreen tree, with smooth bark and very dark green spicy-scented foliage. Bay Rum Birchberry or Privet eugenia - Eugenia ligustrina A pretty shrub or small tree that makes an excellent hedge. The berries are edible, with a sweet, juicy, and slightly aromatic flesh. Black calabash - Amphetheca latifolia A native relative of the commoner calabash or gobi tree, black calabash is restricted to the larger guts and areas that receive a lot of freshwater runoff. -

Morfologia Polínica De Malvaceae: Implicações Taxonômicas E Filogenéticas
MARILEIDE DIAS SABA MORFOLOGIA POLÍNICA DE MALVACEAE: IMPLICAÇÕES TAXONÔMICAS E FILOGENÉTICAS FEIRA DE SANTANA - BAHIA 2007 Livros Grátis http://www.livrosgratis.com.br Milhares de livros grátis para download. UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS -GRADUAÇÃO EM BOTÂNICA MORFOLOGIA POLÍNICA DE MALVACEAE: IMPLICAÇÕES TAXONÔMICAS E FILOGENÉTICAS MARILEIDE DIAS SABA Tese apresentada ao Programa de Pós- Graduação em Botânica da Universidade Estadual de Feira de Santana como parte dos requisitos para a obtenção do título de Doutor em Ciências - Botânica . ORIENTADOR : PROF . DR. FRANCISCO DE ASSIS RIBEIRO DOS SANTOS (UEFS) CO-ORIENTADORA : PROFA . DRA . GERLENI LOPES ESTEVES (IB T/SMA-SP) FEIRA DE SANTANA - BA 2007 BANCA EXAMINADORA ______________________________________________ Profa. Dra. Vânia Gonçalves Lourenço Esteves Museu Nacional - UFRJ _______________________________________________ Prof. Dra. Maria Amélia Vitorino da Cruz Barros Instituto de Botânica (SP) ________________________________________________ Prof. Dra. Cláudia Elena Carneiro Universidade Estadual de Feira de Santana ________________________________________________ Prof. Dr. Luciano Paganucci de Queiroz Universidade Estadual de Feira de Santana _________________________________________________ Prof. Dr. Francisco de Assis Ribeiro dos Santos Orientador e Presidente da Banca - UEFS Feira de Santana - BA 2007 A Deus, toda glória, honra e louvor. Aos amores da minha vida: Jad, Luan e meus pais. Ao mestre e amigo: Francisco, carinhosamente, “Chico”. AGRADECIMENTOS “Você pode sonhar..., criar e construir a idéia mais maravilhosa do mundo, mas são necessárias pessoas para fazer o sonho virar realidade”. (W. Disney) A Deus, que se fez presente em todos os momentos da minha vida, permitindo a conclusão de mais uma etapa. Ao meu orientador, Prof. Dr. Francisco de Assis, pelo incentivo, dedicação e apoio tão importantes para mim. -
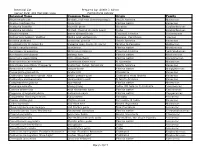
Botanical Name Common Name Origin Family
Botanical List Prepared by: Arielle J. Simon Corner Park: 401 Hampton Lane Horticultural Advisor Botanical Name Common Name Origin Family Abutilon pictum fireball, red vein flowering maple South America Malvaceae Acacia choriophylla cinnecord Florida native Fabaceae Acalypha hispida chenille plant Oceania Euphorbiaceae Acalypha pendula firetail, trailing chenille plant Cuba Euphorbiaceae Aloysia virgata sweet almond bush Tropical America Verbenaceae Anthurium hookeri 'Ruffles' bird's nest anthurium Guyana, Caribbean Araceae Arachis glabrata perennial peanut South America Fabaceae Arachnothryx leucophylla Panama rose, huele de noche Mexico to Panama Rubiaceae Ardisia escallonioides marlberry Florida native Myrsinaceae Asclepias curassavica Mexican milkweed Tropical America Asclepiadaceae Blechnum serrulatum swamp fern Florida native Blechnaceae Bourreria cassinifolia little strongback Florida native Boraginaceae Brachychiton acerifolius Australian flame tree N Australia Malvaceae Brunfelsia pauciflora 'Compacta' yesterday, today, tomorrow South America Fabaceae Byrsonima lucida locust-berry Florida native Malpighiaceae Caesalpinia granadillo bridal veil Venezuela Fabaceae Calliandra haematocephala 'Alba' white powder-puff Cultivated from Bolivia Fabaceae Calliandra surinamensis pink powder-puff N South America Fabaceae Calyptranthes pallens spicewood Florida native Myrtaceae Cananga odorata ylang-ylang India, SE Asia to N Australia Annonaceae Canella winterana wild cinnamon bark Florida native Canellaceae Capparis cynophallophora Jamaican -

Jost Van Dyke, British Virgin Islands
An Environmental Profile of the Island of Jost Van Dyke, British Virgin Islands including Little Jost Van Dyke, Sandy Cay, Green Cay and Sandy Spit This publication was made possible with funding support from: UK Foreign and Commonwealth Office Department for International Development Overseas Territories Environment Programme An Environmental Profile of the Island of Jost Van Dyke, British Virgin Islands including Little Jost Van Dyke, Sandy Cay, Green Cay and Sandy Spit An Initiative of the Jost Van Dykes (BVI) Preservation Society and Island Resources Foundation 2009 This publication was made possible by Use of Profile: Available from: the generous support of the Overseas Reproduction of this publication, or Jost Van Dykes (BVI) Preservation Territories Environment Programme portions of this publication, is Society (OTEP), UK Foreign and authorized for educational or non- Great Harbour Commonwealth Office, under a commercial purposes without prior Jost Van Dykes, VG 1160 contract between OTEP and the Jost permission of the Jost Van Dykes (BVI) British Virgin Islands Van Dykes (BVI) Preservation Society Preservation Society or Island Tel 284.540.0861 (JVDPS), for implementation of a Resources Foundation, provided the www.jvdps.org project identified as: source is fully acknowledged. www.jvdgreen.org BVI503: Jost Van Dyke’s Community- based Programme Advancing Citation: Island Resources Foundation Environmental Protection and Island Resources Foundation and Jost 1718 P Street Northwest, Suite T-4 Sustainable Development. Van Dykes (BVI) Preservation Society Washington, DC 20036 USA (2009). An Environmental Profile of the Tel 202.265.9712 The JVDPS contracted with Island Island of Jost Van Dyke, British Virgin Fax 202.232.0748 Resources Foundation to provide Islands, including Little Jost Van Dyke, www.irf.org technical services as a part of its Sandy Cay, Green Cay and Sandy agreement with OTEP, in particular to Spit.