The Epistemic Revolution Induced by Microbiome Studies: an Interdisciplinary View
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
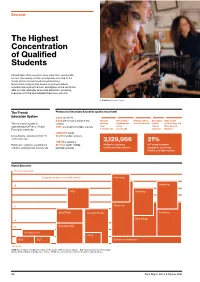
The Highest Concentration of Qualified Students
Education The Highest Concentration of Qualified Students Paris Region offers a world-class education system with an ever-increasing number of programs catering to the needs of international students and families. Universities, engineering, business and specialized schools rank among the most prestigious in the world and offer an internationally renowned education, providing everyone with the best adapted training curriculum. © GOBELINS, l'école de l'image The French Primary and Secondary Education (public and private) Education System 3,200 nurseries 6,925 pre-schools and primary Nursery Pre-school / Primary school Secondary High school The education system is schools starting Kindergarten 6 to 10 years old school 16 to 18 years old operated by the French Public 1,997 secondary and high schools from 2-3 to 11 to 15 Baccalauréat Education Authority. 3 months old 5 years old years old Diploma 1,330,500 pupils Schooling is compulsory from 6 90.2% in public schools to 16 years old. 1,087,184 students 3,129,966 21% Public pre-school is available for 80.7% in public middle students in primary, of France’s student children starting from 2 years old. and high-schools middle and high-schools population in primary, middle, and high schools Higher Education Post A-Level year European grades and credits (ects) Internship Internship +8 PHD Internship +5 Medecine MASTER’S Grandes Ecoles Pharmacy Odontology +3 +2 BACHELOR’S Pro Bachelor’s CGPE BTS DUT Entrance examination A-Levels MESR (French Ministry of Higher Education and Research) - BTS: Technical programs - DUT : University Diploma of Technology Grandes Ecoles : Engineering & Business schools - CPGE : class which prepares students to enter the Grandes Ecoles 20 Paris Region Facts & Figures 2021 Education Highly Educated Students Main Branches of Higher Education (in 2019-20) 91 doctoral schools Bachelor’s, Master’s, and PhD: University programs in technology: 17,566 PhD students, incl. -

Marine Microorganisms: Evolution and Solution to Pollution Fu L Li1, Wang B1,2
COMMENTARY Marine microorganisms: Evolution and solution to pollution Fu L Li1, Wang B1,2 Li FL, Wang B. Marine microorganisms: Evolution and solution to pollution. J Mar Microbiol. 2018;2(1):4-5. nce ocean nurtured life, now she needs our care. Marine microorganism will be an opportunity to further understand ourselves and to seek for new Ois the host of ocean in all ages. We should learn from them humbly. methods of fighting old infections. Marine microorganism is tightly bond with human during the history of evolution and nowadays’ environment pollution. Along with industrial revolution, our marine ecosystem suffered serious pollutions. Microplastics are tiny plastic particles (<5 mm) (Figure 1B), Although the topic is still in debate, life is probably originated from which poison marine lives. Because these microplastics are very hard to be submarine in hydrothermal vent systems (1). In the journey of evolution, our degraded, it is predicted that there will be more microplastics than fish in biosphere was completely dominated by microbes for a very long time (Figure ocean by the year 2050 (7). Since marine sediments are considered as the sink 1A). Human being evolves with those microorganisms. Consequently, of microplastics and marine microbes are key dwellers of marine sediments, the influences of microorganisms can be found in all aspects of human more attention should be paid on the interactions between microplastics biology. More than 65% of our genes originated with bacteria, archaea, and and marine microbes. Actually, a call for this has been published in 2011 unicellular eukaryotes, including those genes responsible for host-microbe (8). -

ASPIRE Exchanges Factsheet Sorbonne University
ASPIRE Exchanges Factsheet Sorbonne University Paris, France Spring Semester 2020 GENERAL INFORMATION University Sorbonne University (SU) Student body 55,600 International students 10,200 at university Main site: http://www.sorbonne-universite.fr/en Exchange site: Sciences and engineering: http://sciences.sorbonne- Website universite.fr/en/international/international_applicants/exchange_progra mme.html Arts and humanities: http://lettres.sorbonne-universite.fr/l-international • Faculty of Science and Engineering Academic units • Faculty of Arts and Humanities Course load IU students are expected to take 15 credits (30 ECTS per semester) Faculty of Science and Engineering: https://www.sorbonne- universite.fr/en/academics/degree-programs/sciences-engineering Course catalogs Faculty of Arts and Humanities: http://vof.paris- sorbonne.fr/fr/index.html French Students should have sufficient oral and written skills in French. SU Language of recommends the B1/B2 (intermediate high) level on the CEFRL. A student instruction who has successfully completed at least one 300-level class is likely to be at this level. Note: Some master’s level classes are conducted in English. Students may consider these on a case-by-case basis and must consult with their academic advisors. https://cts.admissions.indiana.edu/transferin.cfm Course equivalencies (U Paris Sorbonne and U Pierre Marie Curie) SU uses number grades based on a 20-point grading scale, with 10 being the passing point. SU will provide transcripts for students taking part in Grades and the exchange. At the student’s request, the College will review courses to Transcripts determine how they will apply to your degree. Please note that IU does not accept Pass/Fail. -

Phytoplankton As Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate
sustainability Review Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate Samarpita Basu * ID and Katherine R. M. Mackey Earth System Science, University of California Irvine, Irvine, CA 92697, USA; [email protected] * Correspondence: [email protected] Received: 7 January 2018; Accepted: 12 March 2018; Published: 19 March 2018 Abstract: The world’s oceans are a major sink for atmospheric carbon dioxide (CO2). The biological carbon pump plays a vital role in the net transfer of CO2 from the atmosphere to the oceans and then to the sediments, subsequently maintaining atmospheric CO2 at significantly lower levels than would be the case if it did not exist. The efficiency of the biological pump is a function of phytoplankton physiology and community structure, which are in turn governed by the physical and chemical conditions of the ocean. However, only a few studies have focused on the importance of phytoplankton community structure to the biological pump. Because global change is expected to influence carbon and nutrient availability, temperature and light (via stratification), an improved understanding of how phytoplankton community size structure will respond in the future is required to gain insight into the biological pump and the ability of the ocean to act as a long-term sink for atmospheric CO2. This review article aims to explore the potential impacts of predicted changes in global temperature and the carbonate system on phytoplankton cell size, species and elemental composition, so as to shed light on the ability of the biological pump to sequester carbon in the future ocean. -

2018 Booklet (PDF, 1Mo)
EUROPEAN STUDIES PROGRAMME 2018 TABLE OF CONTENTS SCIENCES PO AT A GLANCE .......................................................................................................... 3 PROGRAMME ..................................................................................................................................... 4 The team ........................................................................................................................................... 4 Overview of the programme ............................................................................................................. 7 Course structure ............................................................................................................................... 8 Course syllabus ................................................................................................................................ 9 Recommended readings ................................................................................................................ 11 Course planning .............................................................................................................................. 14 Institutional visits in Paris ............................................................................................................... 15 USEFUL INFORMATION .................................................................................................................. 16 The library ...................................................................................................................................... -

Academic Grades & Notation Sorbonne University
International Relations Office Incoming Erasmus+ Mobility Cécile Rochefort & Roberta Giubrone Phone : 00.33 (0)1.40.46.47.79 [email protected] ACADEMIC GRADES AND NOTATION AT SORBONNE UNIVERSITY FACULTY OF ARTS AND HUMANITIES Sorbonne University is using the European Credit Transfer and Accumulation System (ECTS). The European Credit Transfer and Accumulation System (ECTS) is a tool which enables students to collect credits for learning achieved through higher education. ECTS credits are based on the workload students need in order to achieve expected learning outcomes. At Sorbonne University, all our marks are out of 20. For information, Sorbonne University – Faculty of Arts and Humanities stands amongst one the most difficult grading systems in France. It is important to note that a 10/20 is a good grade, and that it stands for the passing grade. The credit part refers to the number of European credits the course provides. It is based on the 30 credits per semester system that French students must pass to validate one semester. It does not refer to the number of hours taken for one course. When noted “ Absentee ”, it means that your student did not attend the course. When noted “ Attended ”, it means that your student attended the course but didn’t sit the exam. You can freely choose to give them the ECTS or not. When noted “ Passed ”, it means that your student attended the course and passed the exam, but there is no grade because the professor doesn’t give grades to their students. The local grade ( /20) is the most relevant information you should consider to validate your students’ semester/year at Sorbonne University - Faculty of Arts and Humanities . -

Fall Semester 2021)
Sorbonne university : List of courses opened to 4EU+ students (fall semester 2021) Course title Responsible University Description Language Prerequisites Credits Duration Dates Learning content & Type of objectives examination Representation of Lubov Jurgenson Sorbonne The twentieth century has often been thought of French & 2 or 3 Course starts in Literary and artistic The student violence and (luba.jurgenson@wa Université as a century of mass violence, of which Central Russian according to September – currents of the 20th must produce a literary modernity nadoo.fr) Europe and Russia (and later the USSR) were the the work exact date to be century and their personal work on privileged theater. This violence, at the origin of provided by announced dialogue with the one of the profound cultural mutations, had a considerable the student events that provoked seminar themes: influence on the evolution of epistemological Every second mass violence. either a frameworks in literary studies and the humanities Tuesday (3-6 pm) presentation or a and gave rise to a protean corpus of literary, written paper testimonial or theoretical narratives (and even visual works) that question the legitimacy of culture and knowledge while contributing to the construction of new representational models. How does this complexity relate to the literary modernities of the 20th century? Multiculturality in Delphine Bechtel Sorbonne "Museum, monument, memorial site in Central French & 2 or 3 Course starts in Study of the Students will Central and (delphine.bechtel@ -

1 Sandra Rigot Associate Professor Of
Sandra Rigot Associate Professor of Economics at University Sorbonne Paris 13 Born in Paris French Citizenship Research Fellow at Centre d’Economie Paris Nord (CEPN) Research Fellow at Chaire Energy and Prosperity (Ecole Polytechnique – Ecole Normale Supérieure - ENSAE) Research Fellow at IFRIS (Institute For Research and Innovation in Society) University Sorbonne Paris 13 99 boulevard J-B Clément, 93430 Villetaneuse E-Mail: [email protected] Office: +0033 (0)1.49.40.38.39 ACADEMIC EDUCATION 2014 Ability to Supervise PhD Students in Economics (University Sorbonne Paris 13) Title of dissertation: Strategy and Regulation of Financial Intermediaries Related to Long-term Investment Supervisor : Pr. Dominique Plihon (University Sorbonne Paris 13) Examiners : Pr. Michel Aglietta (Paris-Nanterre University) ; Pr. Florence Legros (Paris- Dauphine University), Pr. Hélène Raymond (Paris-Nanterre University) ; Pr. Robert Guttmann (University Sorbonne Paris 13) 2010 PhD in Economics (Paris-Nanterre University) Magna Cum Laude Title of thesis: Strategy and Governance of Pension Funds Doctoral Supervisors: Pr. Michel Aglietta (Paris-Nanterre University) ; Pr. Laurence Scialom (Paris- Nanterre University) Examiners: Christian de Boissieu (PR, Paris-Panthéon Sorbonne University) ; Gunther Capelle-Blancard (Paris-Panthéon Sorbonne University) ; Olivier Garnier (Chief Economist, Société Générale Bank), Pr. Pierre-Yves Gomez (EM Lyon) 2008 Master’s Degree in Business Law (Paris-Panthéon Sorbonne University) 2006 Master’s Degree in Economics (Paris-Nanterre -

The Academic Structures of Boston, London and Paris: a Comparison Report Prepared for CNRS
The academic structures of Boston, London and Paris: a comparison Report prepared for CNRS CLIENT: Centre National de la Recherche Scientifique (CNRS) DATE: October 2016 [email protected] - www.sirisacademic.com 2 THE ACADEMIC STRUCTURES OF BOSTON, LONDON AND PARIS: A COMPARISON October 2016 Submitted to: Centre Nationnal de la Recherche Scientifique By SIRIS Academic Av. Francesc Cambó, 17 08003 Barcelona Spain Tel. +34 93 624 02 28 [email protected] www.sirisacademic.com For more information and comments about this report, please contact us at [email protected] 3 EXECUTIVE SUMMARY ..................................................................................................................................... 6 KEY RECOMMENDATIONS ................................................................................................................................ 8 LINKS TO THE INTERACTIVE VISUALISATIONS PRESENTED IN THIS REPORT .................................................... 12 Rankings overview ..................................................................................................................................................... 12 Maps of science ......................................................................................................................................................... 12 Visualisation of ranking’s qualitative vs. quantitative criteria ................................................................................... 12 Visualisations of ranking’s qualitative criteria vs. -

Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives
marine drugs Review Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives Emiliana Tortorella 1,†, Pietro Tedesco 1,2,†, Fortunato Palma Esposito 1,3,†, Grant Garren January 1,†, Renato Fani 4, Marcel Jaspars 5 and Donatella de Pascale 1,3,* 1 Institute of Protein Biochemistry, National Research Council, I-80131 Naples, Italy; [email protected] (E.T.); [email protected] (P.T.); [email protected] (F.P.E.); [email protected] (G.G.J.) 2 Laboratoire d’Ingénierie des Systèmes Biologiques et des Procédés, INSA, 31400 Toulouse, France 3 Stazione Zoologica “Anthon Dorn”, Villa Comunale, I-80121 Naples, Italy 4 Department of Biology, University of Florence, Sesto Fiorentino, I-50019 Florence, Italy; renato.fani@unifi.it 5 Marine Biodiscovery Centre, Department of Chemistry, University of Aberdeen, Aberdeen, Scotland AB24 3UE, UK; [email protected] * Correspondence: [email protected]; Tel.: +39-0816-132-314; Fax: +39-0816-132-277 † These authors equally contributed to the work. Received: 23 July 2018; Accepted: 27 September 2018; Published: 29 September 2018 Abstract: The increasing emergence of new forms of multidrug resistance among human pathogenic bacteria, coupled with the consequent increase of infectious diseases, urgently requires the discovery and development of novel antimicrobial drugs with new modes of action. Most of the antibiotics currently available on the market were obtained from terrestrial organisms or derived semisynthetically from fermentation products. The isolation of microorganisms from previously unexplored habitats may lead to the discovery of lead structures with antibiotic activity. The deep-sea environment is a unique habitat, and deep-sea microorganisms, because of their adaptation to this extreme environment, have the potential to produce novel secondary metabolites with potent biological activities. -

Marine Microbial Diversity
Marine microbial diversity 27 K. S. Sobhana Marine Biodiversity Division, Central Marine Fisheries Research Institute, Kochi-682 018 Microbes were the only form of life for the first 2-3 billion and also based on iii) concentration of nutrients and required years of planetary and biological evolution. Life most likely growth substances (Oligotrophic, Mesotrophic, Eutrophic). began in the oceans and marine microorganisms are the However, interfaces tend to be hotspots of diversity and closest living descendants of the original forms of life. Early biological activity. Marine microbial habitats at interfaces marine microorganisms also helped create the conditions include the air-water, water-sediment, water-ice, and under which subsequent life developed. More than two billion host macroorganism-water interfaces. The sub-millimeter years ago, the generation of oxygen by photosynthetic marine scale of physical and chemical variability in these habitats microorganisms helped shape the chemical environment in poses a serious challenge to studying interface habitats in which plants, animals, and all other life forms have evolved. detail. The fact that variations can even occur within a few Macroscopic life and planetary habitability completely millimetres, suggests that microbial diversity encompasses depend upon the transformations mediated by complex more than the documented evidence available. Hence, microbial communities. These microscopic factories both biogeography is gaining importance as a field of study from aerobic and anaerobic are the essential catalysts for all of the microbial diversity point of interest. Due to the innately chemical reactions within the biogeochemical cycles. Their small size of the microorganisms, environmental complexity unique metabolisms allow marine microbes to carry out many plays a major role in determining diversity. -

Corals and Sponges Under the Light of the Holobiont Concept: How Microbiomes Underpin Our Understanding of Marine Ecosystems
fmars-08-698853 August 11, 2021 Time: 11:16 # 1 REVIEW published: 16 August 2021 doi: 10.3389/fmars.2021.698853 Corals and Sponges Under the Light of the Holobiont Concept: How Microbiomes Underpin Our Understanding of Marine Ecosystems Chloé Stévenne*†, Maud Micha*†, Jean-Christophe Plumier and Stéphane Roberty InBioS – Animal Physiology and Ecophysiology, Department of Biology, Ecology & Evolution, University of Liège, Liège, Belgium In the past 20 years, a new concept has slowly emerged and expanded to various domains of marine biology research: the holobiont. A holobiont describes the consortium formed by a eukaryotic host and its associated microorganisms including Edited by: bacteria, archaea, protists, microalgae, fungi, and viruses. From coral reefs to the Viola Liebich, deep-sea, symbiotic relationships and host–microbiome interactions are omnipresent Bremen Society for Natural Sciences, and central to the health of marine ecosystems. Studying marine organisms under Germany the light of the holobiont is a new paradigm that impacts many aspects of marine Reviewed by: Carlotta Nonnis Marzano, sciences. This approach is an innovative way of understanding the complex functioning University of Bari Aldo Moro, Italy of marine organisms, their evolution, their ecological roles within their ecosystems, and Maria Pia Miglietta, Texas A&M University at Galveston, their adaptation to face environmental changes. This review offers a broad insight into United States key concepts of holobiont studies and into the current knowledge of marine model *Correspondence: holobionts. Firstly, the history of the holobiont concept and the expansion of its use Chloé Stévenne from evolutionary sciences to other fields of marine biology will be discussed.