Genes, Culture, and Agriculture: an Example of Human Niche Construction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
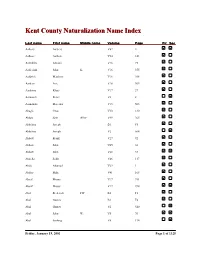
Kent County Naturalization Name Index, Aalbers, A
Kent County Naturalization Name Index Last name First name Middle name Volume Page Fir Sec Aalbers Aalbers V62 4 Aalbers Aalbert V24 141 Aalddriks Antonie V16 75 Aalderink John K. V16 355 Aaldrick Matthew V16 308 Aardem Arie V16 304 Aardema Klaas V17 27 Aarnouds Pieter V6 8 Aarnoudse Marenus V15 503 Abagis Chas V30 130 Abbas Sain Allez V49 265 Abbelma Joseph B1 F5 Abbelma Joseph V2 564 Abbott Frank V27 92 Abbott John V45 36 Abbott John V68 33 Abdella Salik V46 117 Abdo Ahamad V29 1 Abdoo Mike V41 168 Abeaf Moses V17 391 Abeaf Moses V17 394 Abel Frederick FW B1 F1 Abel Gustav B1 F4 Abel Gustav V2 540 Abel John W. V5 70 Abel Ludwig V8 134 Friday, January 19, 2001 Page 1 of 1325 Last name First name Middle name Volume Page Fir Sec Abella Salih V68 85 Abezi Albert V25 76 Aboabsee Theab V74 40 Aboasee Theab V18 150 Abood N. B1 F5 Abood Nemy V3 90 Abraham John V17 381 Abrahamson Charles Y. B7 200 Abram John B1 F1 Abramson Morris B1 F3 Abraursz Abram Peter V27 159 Abromaitis Louis V27 381 Abromaitis Louis V67 90 Absmaier Carl V77 4 Accardi Guiseppe V50 79 Acheson John V16 616 Achille Minciotti V51 142 Achtenhof Jakob V15 145 Achter Jan V17 200 Achterhof Henri B1 F1 Achterhof Johannes V15 500 Achterhof Matheus B1 F1 Achtjes John B7 107 Ackermann Joseph V15 282 Acton John C. B7 222 Adair David G. V15 335 Adair Joseph V15 335 Adalphson Emil V18 197 Friday, January 19, 2001 Page 2 of 1325 Last name First name Middle name Volume Page Fir Sec Adam Frickartz Heinrich V41 279 Adama Jelle V22 176 Adamawiczus Baltris V37 155 Adamczak Peter V38 245 Adamczyk Wladyslaw V35 291 Adams Edward John V24 70 Adams Frank B1 F2 Adams George W. -

4 F\Ihli«N%D by the I Llhunntan Catholu* O Ra Ugi Ja Pro** Aoctt'u' 23.H So Ftakl*; A**
? MlOftEST 1. v£# 5721 COTTAGE ^-U1"1- CttlCAGd 57. 1LL. (JRATIS JUM D fc A U O A S t) H A tr O A S L^idSia Ueiuviy Katalike Iffta 1 » Tft. I ItliiumtaJt Daily Vtim4 f\ihli«n%d by the I llhunntan CatholU* O ra ugi Ja Pro** Aoctt'U' 23.H So ftakl*; A**. < Mo**., •. llllnoU 2231 Sn. Oaktry Are., CMcagn t. Kilnote T r teplione — \ Irglnla 7-A04O •41 N Telepttooo — Vlrfrinla 7-«4*0-4| Vienintelis tautinė* Ir tikybine* mlntl»s The tno«t lnflueatlal I.ithuanlan Daily h*imv%ų llenraitla pasaulyje THE LITHUANIAN DAILY, FRIEND tn America No. 232 Kaina 5 oeatal PIRMADIENIS, SPALIO (OCTOBER) 4, 1954 Priee 5 cente VOL. XXXVII • .-.. .. LONDONO KONFERENCIJOJE SUSITARTA i Formoza Peipingo žaidime Šį mėnesį rinksis dar keliose Indijos premjerą Nchru nepaprastai jaudina susišaudymai, vietose nutarimams tvarkyti kurie vyksta Quemoy — Amoy erdvėje tarp abiejų Kinijos vy riausybių karinių pajėgų. Nehru jaudinasi ne be pagrindo, nes jis, turbūt, goriausiai yra painformuotas apie Peipingo nuotai LONDON, spalio 4. — Devynių valstybių konferencija čia kas. baigėsi vakar pilnu susitarimu dėl V. Vokietijos apginklavimo, jai suverenumo grąžinimo ir jos įjungimo į Briuselio ir Atlanto Galop ir žinomi faktai nėra ~ '. ', raminą. Raudonoji Kinija ima Sią Savaitę SlltartlS paktus. telkti prieš Formozą karines Vakar susitarimai pasirašyti viena iš specialių JT organiza- ne kaip formalios sutartys, bet pajėgas (aviaciją ir parašiuti Triesto ginčui baigti . • . ,J... cijų, tačiau žinoma, kad jos yra kaip principų ir jų praktiško . :. ' ,J . ninkus), o JAV laivynas, dabar labai autonomiškosv , o atominės TRIESTE, sp. 4. — Po devy pritaikymo deklaracijos-proto- žymiai sustiprintas, plaukioja energijos organizacija tos auto nių mėnesių derybų Romoje, kolaj. -

Darbininkas
boston public LIERARY C -1 F C F L * - - - L -■ * 1 ■ ' :•. i.division COPLEY SQ BOSTON MASS 16 3 tUlUUUUlUUUtlUlltUlUUiiimittiUmilUlllliliia 4--------------------------------------------- 4< I DABARTIES DARBININKAS Lithuanian Semi-Weekly < • :j BILDESIIJOS i Newspaper , Published every , Ua. DARBININKAS TUESDAY and FRIDAY J 366 Broaway, S. Boston 27, Mau. 1 DĖMESIUI! AMERIKOS LIE ' K. ŠVENTO JUOZAPO DARBININKŲ SĄJUNGOS ORGANAS ♦---------------------------------------- • < _________ I Kitą savaitę penktadie- VOL. XXX — No. 76. TEL. SOUth Boston 2680 ANTRADIENIS (Tuesday) SPALIŲ (October) 9 D., 1945 M. TEL. SOUth Boston 2680 FIVE CENTS nio “Darbininko” numeris išeis LDS ir “Darbininko” 3,000 Pagerbė Šv. Pran 30-ties metų gyvavimo su kakties paminėjimui. Ta ciškų Asizietį Rusija Pyksta Ant J. V., Britani me numeryj tilps įvairių straipsnių apie LDS ir I Boston, Mass. — Sekma-; “Darbininką” ir vaizdų. dieni, spalių 7 d. trys tūks jos Dėl Japonijos Ir Rytinės tančiai Trečiojo Ordeno— I Gerb. LDS ir “Darbinin Šv. Pranciškaus AsiziečioI. ko” įkūrėjai, bendradar narių susirinko pagerbti ir! biai, LDS apskričiai ir pagarbinti įžymųjį šven-: kuopos kviečiamos Uitąjįp~ į Šv. Kryžiaus Kated Stalinas Nevažiuosiąs Iš Sovietų Rusi puošti tą “Darbininko” nu rą. merį savo sveikinimais ir J. E. arkivyskupas Ri- jos Į Trijų Didžiųjų Susirinkimą atsiminimais. Visus raš chard J. Cushing, D. D., tus prašome siųsti taip, pasakė turiningą pamoks Londonas, spalių 8 Kaip žinoma, sovietų Ru- kad Jūsų redakcija juos lą apie Šv. Pranciškų, jo Santykiai tarp didžiųjų są-J sijos propagandos mašina gautų spalių 16 d. arba gyvenimą ir darbus. jungininkų — sovietų Ru-! atsargiai skleidė idėją ne prieš tą dieną. Vėliau gau į Kun. Pranciškus Juškai sijos iš vienos pusės ir, tik Rusijoje, bet ir kaimy- ti sveikinimai ir straips tis, Trečiojo Ordeno dvasi- Britanijos ir Jung. -

SAE 2006 World Congress & Exhibition
SAE 2006 World Congress & Exhibition Technical Session Schedule As of 04/09/2006 07:40 pm Monday, April 3 Can Racing Partnerships Change to Accelerate Innovation (Focus on Sports Car Racing) Session Code: ANNUAL108 Room FEV Powertrain Innovation Forum Session Time: 10:00 a.m. Professional Sanctioning Organizations Organizers - Herbert A. Fishel Moderators - Paul F. Pfanner, Founder, Racer Communications Inc. Panelists - James Julow, President, Sports Car Club of America; H. Doug Robinson, Executive Director, International Motor Sports Association; Scott Atherton, President & CEO, American Le Mans Series; Roger Edmondson, President, Grand American Road Racing; Stephane Ratel, CEO, SRO, Ltd. Monday, April 3 Can Racing Partnerships Change to Accelerate Innovation (Focus on Sports Car Racing) Session Code: ANNUAL109 Room FEV Powertrain Innovation Forum Session Time: 2:00 p.m. Automobile Manufacturers Organizers - Herbert A. Fishel Moderators - John McElroy, President, Blue Sky Productions Panelists - Richard T. Brekus, Mgr, Product Planning & Strategy, BMW of North America; John M. Doonan, Mgr, Motorsports Team Dev, Mazda North American Operations; Marcus Haselgrove, Team Manager, Technical Director, B-K Motorsports Inc.; David R. Wilson, TRD Group VP-Strategic Planning, Fin & Ops, Toyota; Steven J. Wesoloski, Road Racing Group Manager, GM Racing, General Motors Corp.; Robert Davis, Senior VP, Quality R&D, Mazda North American Operations; Frank-Steffen Walliser, General Manager Motorsport Strategy, Porsche AG Monday, April 3 Grand Opening Session Code: ANNUAL100 Room AVL Technology Theater (open to all Session Time: 9:30 a.m. Keynote Speakers - Remarks by: Burkhard Goeschel, Member of the Board of Mgmt., BMW AG Develop and Purchasing; Richard O. Schaum, SAE Automotive Vice President; Helmut List, President & CEO, AVL LIST GmbH Monday, April 3 Innovation in the Automotive Industry - An Extended Session Covering Managing Innovation and Driving Innovation with Research Session Code: ANNUAL101 Room AVL Technology Theater (open to all Session Time: 10:00 a.m. -

Kundenmagazin "IN the LEAD" 5
LANDESHAUPTSTADT RHEINMAIN CONGRESSCENTER WIESBADEN MAG 05| 2018 INTHE FUTURE OFTHE CONGRESS AND CONVENTION LEAD VISION ERFÜLLT DAS „INNOVATIVSTE KONGRESS- UND VERANSTALTUNGSZENTRUM DEUTSCHLANDS“ Vision fulfilled – Germany’s most innovative convention and event centre 13. UND 14. APRIL 2018 GRAND OPENING 13th and 14th April 2018 – Grand opening EINSATZ, ENGAGEMENT UND LEIDENSCHAFT EIN HAUS SAGT DANKE Commitment, dedication and passion – a building says thank you HENNING WOSSIDLO „WIR HABEN DIE CHANCE GENUTZT“ Henning Wossidlo – “We seized the opportunity” www.wiesbaden.de CONTENT S. 05 S. 11 S. 18 + SERVICE + DESIGN + BACKGROUND 11 Teamgeist TEAM SPIRIT GESCHAFFT! 04 Vision erfüllt 18 Kinderleicht A FAIRY TALE OF 1001 SIMPLE OPTIONS VISION FULFILLED 07 Mission Zukunft RheinMain CongressCenter S. 08 MISSION FUTURE + CLIENTS 22 Global Greetings + PERSONALITY S. 12 GLOBAL GREETINGS + TRENDS + LIVE S. 21 08 Grand Opening A LAVISH CELEBRATION FOR THE WHOLE OF WIESBADEN 10 Machen Sie den Wohlfühl-Check MAKE THE FEEL-GOOD CHECK 12 Wossidlo, Pflugradt: „Wir haben die Chance genutzt“ 15 Barrierefreie Events und Kongresse 21 RMCC erleben “WE SEIZED THE OPPORTUNITY“ BARRIER-FREE EVENTS AND CONGRESSES EXPERIENCE RMCC 16 Ein Haus sagt Danke 20 Digitale Zeichensprache 26 Auf ein letztes Wort, Comic A BUILDING SAYS THANK YOU DIGITAL SIGNAGE IN THE RMCC FINAL WORDS, COMIC LIEBE LESERINNEN UND LESER, DEAR READERS — Home straight: The curtain goes up from 13th to 14th April. Experience the great opening weekend. das RheinMain CongressCenter befindet sich auf der finalen Zielgeraden: Vom 13. bis 14. April geht der Vor- We spent 36 weeks working on realising the major new construction project. hang auf. Erleben Sie das große Eröffnungswochenende, feiern Sie mit uns den Beginn einer neuen Ära im A successful mammoth project thanks to an excellent performance. -

A and No Surname
-A- Surname Index and without a surname St. Clair County Genealogical Society Quarterly Volumes 1–40 (1978-2017) Nuns are indexed under “S” as Sister [given name] To find all instances of your family name, search for variants caused by poor handwriting, misinterpretation of similar letters or their sounds. A few such examples are L for S, c for e, n for u, u for a; phonetic spellings (Aubuchon for Oubuchon); abbreviations (M’ for Mc); single letters for double (m for mm, n for nn); translations (King for Roy, Carpenter for Zimmermann). Other search tips: substitute each vowel for other ones, search for nicknames, when hyphenated – search for each surname alone, with and without “de” or “von”; with and without a space or apostrophe (Lachance and La Chance, O’Brien and OBRIEN). More suggestions are on the SCCGS website Quarterly pages at https://stclair-ilgs.org . Surname Vol. Issue Page(s) Surname Vol. Issue Page(s) Surname Vol. Issue Page(s) ___ , Cunagunda Isabella27 2 81 ___, George 24 3 108 ___, unknown 34 2 87, 89, 91, 93, ___ , Eugene 33 2 62 ___, George H. 24 3 111 96, 101 ___ , female child 33 3 135 ___, Illegible 27 1 32, 37 ___, unknown 34 3 150, 156 ___ , female infant 30 4 217 ___, Illia 24 3 116 ___, unknown 35 2 63, 73, 77 ___ , female newborn 33 2 65, 69 ___, INDIANS 16 2 109 ___, unknown 35 2 69, 70, 76 ___ , female, 25y 31 2 65 ___, Infant 38 1 45 ___, unknown 35 3 161, 162, 154 ___ , female, age 32 31 1 29 ___, Infant 38 2 92 ___, unknown 36 1 95, 96, 98, 110 ___ , Jacob 26 2 98 ___, Jacco 16 2 91 ___, unknown 36 3 122 -

Our Affiliates
OUR AFFILIATES CONTINENT COUNTRY SURNAME/NAME Africa Congo Gupa Makila Marcellin Africa Kenya Mulindi Victor Africa Mauritius Bhoyrul Dustin Africa Morocco Berrada Meriem Africa Morocco Alassaire Sylvain Africa Namibia Samuel Alvine Mirjam Africa Nigeria Olufunmilayo “Funmi” Oju Africa Nigeria Sanda Akin Africa Nigeria Nwaokobia Nnenna Africa South Africa Biseswar Kirthi Africa South Africa Fourie Wynand Africa Uganda Ogwang Tommy Africa Uganda Mbabaali Mariam Africa Uganda Kentaro Mugerwa Eva Africa Uganda Tibugwisa Damalie Africa Zimbabwe Matsanura Paddington Asia China Che Michelle Asia Hong Kong Law Elsa Asia Hong Kong Fung Kin Wah Franky Asia India Divekar Pallavi Asia India Merchant Dimple Asia India Bansal Kushagra Asia India Ganu Sayali Asia India Bansal Rashi Asia Iran Shariatpanahy Sahar Asia Israel Weinrauch Joseph Asia Japan Mine Masaki Asia Japan Kikegawa Takumi Asia Japan Matsuzawa Koh Asia Japan Nakamura Yuki Asia Japan Watanabe Masashi Asia Japan Usui Yoshinori Asia Japan Gemba Kazuko Asia Japan Moriyama Toshio Asia Japan Hara Takashi Asia Malaysia Cheah Joyi Asia Malaysia Nik Erman Nik Roseli willeague.com OUR AFFILIATES CONTINENT COUNTRY SURNAME/NAME Asia Malaysia Sinappan Sathya Asia Malaysia Zainal Anwar Ezzad Asia Malaysia Sim Wen Yee Asia Philippines Clet Nestie Asia Philippines Doligon Daniel Asia Singapore Chong Yue-En Asia Singapore Tan Mark Asia South Korea Shin Mooyun Asia Taiwan Chen Donna Asia Taiwan Yang Elvy Asia Thailand Suthisarnsuntorn Wayu Asia Un. Arab Emirates Schildgen Elena Angelika Asia Un. Arab -

Annual Report
coast guard foundation 2015 annual report coast guard foundation z message from leadership 2015 highlights invite you to take a look at the stories and photos • Awarded 86 new scholarships valued at on these pages — you’ll see an amazing group of more than $400,000 to children of enlisted people who have dedicated their lives to serving Coast Guard members. This includes three I Fallen Heroes Scholarships to children of others, caring for our environment, ensuring our Coast Guard heroes who have died in the line safety on the water, protecting our waterways and of duty. Our three “Sara Statts” scholars will ports, stopping dangerous drugs from making their have 100 percent of their college expenses way into our neighborhoods, and so much more. covered by the Coast Guard Foundation. You’ll also learn about some of our generous support- • Distributed more than $388,000 in ers who have made significant and impactful gifts scholarships and grants for active duty enlisted Coast Guard members who are that make a real difference for Coast Guard members and their families. pursuing college or advanced degrees I’m hard-pressed to say what has made the biggest impression on me while serving in the Coast Guard. this year at the Coast Guard Foundation. Our chairman of the board, Bob • Awarded $60,000 in grants and Flynn, passed away in February of 2015 and in his memory, we established scholarships for spouses of active duty a fund to support the recruitment and retention of underserved popula- enlisted Coast Guard members, our tions at the United States Coast Guard Academy, an initiative that was near largest single award-year yet. -

Carlson Crewmember Only
A‐B 11/10/2009 STATE OF ALASKA Commercial Fisheries Entry Commission Carlson Summary Crewmember License Holders ONLY Obs Name Obs Name Obs Name 1 AADNES, STEWART W. 2 AADSEN, TELE L. 3 AAFF, DANIEL L. 4 AAGAARD, PETER G. 5 AAKER, JESSIE J. 6 AAL, MARTIN A. 7 AALAND, DAVID R. 8 AALAND, RICHARD D. 9 AALCOMB, CHUCK J. 10 AALONA, ALLAN J. 11 AAMOT, JAN E. 12 AANDENID, MARK 13 AANDERUD, ZACHARY T. 14 AAREN, RUSSELL P. 15 AARHAUG, TORFINN 16 AARON, DARWIN A. 17 AARON, JONATHAN G. 18 AARON, WAYNE R. 19 AARON, WILLIAM H. 20 AARON, WILLIAM O. 21 AASE, ANGELA M. 22 AASE, HENNING 23 AASEN, JON J. 24 AASNESS, JOHN C. 25 ABAD, JOSEPH C. 26 ABADIE, BERNARD 27 ABADY, MOHAMMED E. 28 ABALAMA, SHANNON R. 29 ABALATEO, RENATO S. 30 ABALOS, ANTHONY 31 ABALOS, HERMAN G. 32 ABAN MOO, WILLIAM F. 33 ABAN, FREDDY T. 34 ABATE, TIM D. 35 ABBE, THOMAS E. 36 ABBEY, CHRIS W. 37 ABBEY, STEVEN C. 38 ABBEY, WAYNE T. 39 ABBOTT, ADAM J. 40 ABBOTT, BRUCE K. 41 ABBOTT, BRYAN 42 ABBOTT, CARLOS F. 43 ABBOTT, CHARLES V. 44 ABBOTT, DAVID N. 45 ABBOTT, DENNIS R. 46 ABBOTT, ERIC S. 47 ABBOTT, GREG P. 48 ABBOTT, HANS T. 49 ABBOTT, HAROLD W. 50 ABBOTT, JACK T. 51 ABBOTT, JIM 52 ABBOTT, JOHN P. 53 ABBOTT, MARK A. 54 ABBOTT, ROBERT L. 55 ABBOTT, ROBERT W. 56 ABBOTT, ROSS W. 57 ABBOTT, ROY W. 58 ABBOTT, WAYNE T. 59 ABBOTTMOORE, MATTHEW A. 60 ABDAL‐SHAHEED, NASIF M. 61 ABDELHADI, ADAM T. -

Forum on Physics and Society
1 Forum on Physics and Society TM Definition: A name preceded by an asterisk (*) denotes that individual as a Fellow; a double dagger (‡) means that a person is a lifetime member. All other names are regular members. A al-Binni, Usama A ancalle, liian a Artz, Jerry L Bacher, Andrew Aastuen, David J W Al-Hagan, Ola Andeen, Timothy R Asai, Kichizo Bacon, Trevor C ‡Abbas, Daniel C al-Wahish, Amal B Andelin, John P Asakawa, Masayuki Bacrania, Minesh K Abbott, Anthony J Alban, Eduardo Xavier Anderson, Christopher P Asaro, Catherine Ann Baddour, Nicolas Abbott, Heather L. Albayrak, Elif Asli ‡Anderson, Gordon Wood Ashpis, David E Badu, Shyam Raj Abdel-Naby, Shahin Albayrak, Ibrahim H *Anderson, J E Ashton, Stephen Ernest Baer, Donald R Ahmed Albers, David J Anderson, Kathleen Askenazy, Philip D Bagaipo, Jupiter Anthony Abel, George K Albright, Brian James ‡Anderson, Leonard Asmat, Martin Antonio G *Abella, Isaac D *Albright, David Mahlon Aspelmeyer, Markus ‡*Bagger, Jonathan A. Aberg, Denny K T Alduhaileb, Raed A Anderson, Matthew P Aspuru-Guzik, Alan ‡Baggett, Neil V Abeshouse, David J Aleinikava, Darya Anderson, Michael G Aston, David Baglin, Coral M Abeyta, Adam C Alem, Orang *Anderson, Philip W Asuquo, Cletus Baglin, Frank G Abeyweera, Buddika K Aleman, Benjamin J Andino, Richard S Asztalos, Stephen John Bai, Xianming Abi-Salloum, Tony ‡Alexander, Michael N Andresen, Kurt Atalay, Funda Bai, Zhifeng Youssef Alexander, Paris Andrews, James Herbert Ates, Ozgur *Baierlein, Ralph F *Abkowitz, Martin A Alexander, Steven A Animalu, Alexander O.E. Atkins, Scott Bailey, Catherine Noel Abolfathi, Bela *Alexeff, Igor Animilli, ShravanRakesh Atkinson, Juan Miguel Baillie, Jeannette M. -

Genetic Analysis of Uniparental and Autosomal Markers in Human Populations
UNIVERSIDADE DE SANTIAGO DE COMPOSTELA FACULTADE DE MEDICINA E ODONTOLOXÍA INSTITUTO DE CIENCIAS FORENSES “LUÍS CONCHEIRO” Genetic analysis of uniparental and autosomal markers in human populations Memoria que presenta, para optar al grado de doctor, Francesca Brisighelli Santiago de Compostela, 2012 El Doctor D. Ángel Carracedo Álvarez, Catedrático de Medicina Legal de la Facultad de Medicina de la Universidad de Santiago de Compostela, y el Doctor D. Antonio Salas Ellacuriaga, Profesor Titular en la Universidad de Santiago de Compostela, CERTIFICAN: Que la presente memoria que lleva por título “Genetic analysis of uniparental and autosomal markers in human populations”, de la licenciada en Sciencias Naturales por la Universidad de Roma “La Sapienza” Francesca Brisighelli, ha sido realizada bajo nuestra dirección, considerándola en condiciones para optar al Grado de Doctor y autorizándola para su presentación ante el Tribunal correspondiente. Y para que así conste, firmamos la presente en Santiago de Compostela, . Prof. Dr. Ángel Carracedo Álvarez Dr. Antonio Salas Ellacuriaga Francesca Brisighelli ai miei genitori Ringraziamenti ….e’ proprio vero, solo adesso che mi ritrovo di fronte questo foglio bianco, mi rendo conto che scrivere questa parte non è affatto facile! Spero di non dimenticare nessuna delle persone che mi hanno accompagnato in questo meraviglioso percorso, non solo dal punto di vista lavorativo, ma anche, e soprattutto, di crescita personale. Iniziamo…. Grazie, a Ángel, per aver creduto in me, dandomi la possibilità di unirmi al tuo gruppo di lavoro e poter così anche sperimentare ciò che significa vivere fuori dal proprio paese a Vincenzo, per avermi, in quel momento, “lasciato andare” dal laboratorio di Roma, incitandomi ad iniziare questa impegnativa ma incredibile esperienza a Maviky, per l’appoggio incondizionato, per le chiacchiere e per i consigli, per ciò che riguarda il lavoro ma anche il resto a Toño, “el Xefiño”: mi risulta difficile riassumere in 2 righe la gratitudine che ti devo. -

Anllech an Aolt an Boer an C an Cash an Caskey An
BUSCAPRONTA www.buscapronta.com ARQUIVO 08 DE PESQUISAS GENEALÓGICAS 61 PÁGINAS – MÉDIA DE 19.600 SOBRENOMES/OCORRÊNCIA Para pesquisar, utilize a ferramenta EDITAR/LOCALIZAR do WORD. A cada vez que você clicar ENTER e aparecer o sobrenome pesquisado GRIFADO (FUNDO PRETO) corresponderá um endereço Internet correspondente que foi pesquisado por nossa equipe. Ao solicitar seus endereços de acesso Internet, informe o SOBRENOME PESQUISADO, o número do ARQUIVO BUSCAPRONTA DIV ou BUSCAPRONTA GEN correspondente e o número de vezes em que encontrou o SOBRENOME PESQUISADO. Número eventualmente existente à direita do sobrenome (e na mesma linha) indica número de pessoas com aquele sobrenome cujas informações genealógicas são apresentadas. O valor de cada endereço Internet solicitado está em nosso site www.buscapronta.com . Para dados especificamente de registros gerais pesquise nos arquivos BUSCAPRONTA DIV. ATENÇÃO: Quando pesquisar em nossos arquivos, ao digitar o sobrenome procurado, faça- o, sempre que julgar necessário, COM E SEM os acentos agudo, grave, circunflexo, crase, til e trema. Sobrenomes com (ç) cedilha, digite também somente com (c) ou com dois esses (ss). Sobrenomes com dois esses (ss), digite com somente um esse (s) e com (ç). (ZZ) digite, também (Z) e vice-versa. (LL) digite, também (L) e vice-versa. Van Wolfgang – pesquise Wolfgang (faça o mesmo com outros complementos: Van der, De la etc) Sobrenomes compostos ( Mendes Caldeira) pesquise separadamente: MENDES e depois CALDEIRA. Tendo dificuldade com caracter Ø HAMMERSHØY – pesquise HAMMERSH HØJBJERG – pesquise JBJERG BUSCAPRONTA não reproduz dados genealógicos das pessoas, sendo necessário acessar os documentos Internet correspondentes para obter tais dados e informações. DESEJAMOS PLENO SUCESSO EM SUA PESQUISA.